
It seems every day brings a new scientific refutation of creationism that the frauds who lead the cult and feed it disinformation need to keep hidden, or at least give the half-baked strategies for ignoring. The scientists are lying; the scientists get their dates wrong; no matter what the evidence shows, God's holy word trumps it; if you weren't there to see it, it probably didn't happen.
Maybe creationists can be satisfied with such obvious logical fallacies and evidence free assertions, but normal people will allow the evidence to lead their opinions, not the misrepresentations and lies of frauds who know nothing of the subject.
And what better evidence that the world wasn't created out of nothing by magic in a few days about 10,000 years ago, than yet more evidence that there were creatures like fish living on Earth 380 million years before creationist mythology says the magic creation happened; creatures like this fossil air-breathing, lobe-finned fish found in Devonian rocks in Central Australia.
And to make matters worse for creationists, this fish was a close relative of one of the lobe-finned fish that evolved into the terrestrial tetrapods from which all terrestrial vertebrates are descended. Taxonomists have now named this fish, Harajicadectes zhumini.
What information do you have on the Devonian Rocks of Central Australia and how their age was estimated? The Devonian rocks of central Australia are significant geological formations that provide insights into the Earth's history during the Devonian Period, which occurred approximately 419 to 359 million years ago. These rocks are found in various regions across central Australia, including parts of the Northern Territory, Queensland, and South Australia. The age of these Devonian rocks has been estimated through various geological dating methods, including radiometric dating techniques such as uranium-lead dating, potassium-argon dating, and rubidium-strontium dating. These methods rely on the decay of radioactive isotopes within minerals found in the rocks to determine their age. By analyzing the ratios of parent and daughter isotopes in these minerals, scientists can calculate the age of the rocks. In addition to radiometric dating, stratigraphic analysis and fossil evidence have also been used to determine the age of Devonian rocks. Stratigraphic analysis involves studying the layers or strata of rock formations and correlating them with known geological time scales. Fossil evidence found within the rocks, such as plant remains, microfossils, and the remains of ancient marine life, can also provide important clues about the age of the formations. Overall, the age of Devonian rocks in central Australia has been estimated through a combination of radiometric dating, stratigraphic analysis, and fossil evidence, providing valuable insights into the geological history of the region and the evolution of life on Earth during the Devonian Period.And there's that dreaded Uranium-lead (U-Pb) dating that creationists are coming to fear because it is so deadly accurate and impossible to falsify, except by pretending the weak and strong nuclear forces were so weak 10,000 years ago that the half-life of the radioactive isotopes of Uranium used in the dating was short enough then to make 10,000 years look like 360 million years. This would have meant that atoms couldn't have existed when creationists claim a supernatural magician was creating living organism out of them because the forces that bind the nucleus together wouldn't have been strong enough.
The palaeontologists from Flinders University, Australia, Brian Choo, Postdoctoral fellow in vertebrate palaeontology; Alice Clement, Research Associate in the College of Science and Engineering, and Professor John Long, Strategic Professor in Palaeontology, who named and described this fish have published their findings, open access, in the Journal of Vertebrate Paleontology and describe their work and its significance in an article in The Conversation. Their article is reprinted here under a Creative Commons license, reformatted for stylistic consistency:
380-million-year-old predatory fish from Central Australia is finally named after decades of digging
Harajicadectes cruises through the ancient rivers of central Australia ~385 million years ago.
Credit: Brian Choo
Brian Choo, Flinders University; Alice Clement, Flinders University, and John Long, Flinders University
More than 380 million years ago, a sleek, air-breathing predatory fish patrolled the rivers of central Australia. Today, the sediments of those rivers are outcrops of red sandstone in the remote outback.
Our new paper, published in the Journal of Vertebrate Paleontology, describes the fossils of this fish, which we have named Harajicadectes zhumini.
Known from at least 17 fossil specimens, Harajicadectes is the first reasonably complete bony fish found from Devonian rocks in central Australia. It has also proven to be a most unusual animal.
Meet the biter
The name means “Min Zhu’s Harajica-biter”, after the location where its fossils were found, its presumed predatory habits, and in honour of eminent Chinese palaeontologist Min Zhu, who has made many contributions to early vertebrate research.
Harajicadectes was a fish in the Tetrapodomorpha group. This group had strongly built paired fins and usually only a single pair of external nostrils.
Tetrapodomorph fish from the Devonian period (359–419 million years ago) have long been of great interest to science. They include the forerunners of modern tetrapods – animals with backbones and limbs such as amphibians, reptiles, birds and mammals.
For example, recent fossil discoveries show fingers and toes arose in this group.
Devonian fossil sites in northwestern and eastern Australia have produced many spectacular discoveries of early tetrapodomorphs.
But until our discovery, the poorly sampled interior of the continent had only offered tantalising fossil fragments.
A long road to discovery
Our species description is the culmination of 50 years of tireless exploration and research.
Palaeontologist Gavin Young from the Australian National University made the initial discoveries in 1973 while exploring the Middle-Late Devonian Harajica Sandstone on Luritja/Arrernte country, more than 150 kilometres west of Alice Springs (Mparntwe).
Packed within red sandstone blocks on a remote hilltop were hundreds of fossil fishes. The vast majority of them were small Bothriolepis – a type of widespread prehistoric fish known as a placoderm, covered in box-like armour.
Scattered among them were fragments of other fishes. These included a lungfish known as Harajicadipterus youngi, named in honour of Gavin Young and his years of work on material from Harajica.
There were also spines from acanthodians (small, vaguely shark-like fish), the plates of phyllolepids (extremely flat placoderms) and, most intriguingly, jaw fragments of a previously unknown tetrapodomorph.

The moment of discovery when we found a complete fossil of Harajicadectes in 2016. Flinders University palaeontologists John Long (centre), Brian Choo (right) and Alice Clement (left) with ANU palaeontologist Gavin Young (top left).
Credit: Author provided
There were early attempts at figuring out the species, but this proved troublesome. Then, our Flinders University expedition to the site in 2016 yielded the first almost complete fossil of this animal.
This beautiful specimen demonstrated that all the isolated bits and pieces collected over the years belonged to a single new type of fish. It is now in the collections of the Museum and Art Gallery of the Northern Territory, serving as the type specimen of Harajicadectes.

The type specimen of Harajicadectes discovered in 2016.
Credit: Author provided
Up to 40 centimetres long, Harajicadectes is the biggest fish found in the Harajica rocks. Likely the top predator of those ancient rivers, its big mouth was lined with closely-packed sharp teeth alongside larger, widely spaced triangular fangs.
It seems to have combined anatomical traits from different tetrapodomorph lineages via convergent evolution (when different creatures evolve similar features independently). An example of this are the patterns of bones in its skull and scales. Exactly where it sits among its closest relatives is difficult to resolve.

Artist’s reconstruction of Harajicadectes menacing a pair of armoured Bothriolepis.
Credit: Artist: Brian Choo
Similar giant spiracles also appear in Gogonasus, a marine tetrapodomorph from the famous Late Devonian Gogo Formation of Western Australia. (It doesn’t appear to be an immediate relative of Harajicadectes.)
They are also seen in the unrelated Pickeringius, an early ray-finned fish that was also at Gogo.
The earliest air-breathers?
Other Devonian animals that sported such spiracles were the famous elpistostegalians – freshwater tetrapodomorphs from the Northern Hemisphere such as Elpistostege and Tiktaalik.
These animals were extremely close to the ancestry of limbed vertebrates. So, enlarged spiracles seem to have arisen independently in at least four separate lineages of Devonian fishes.
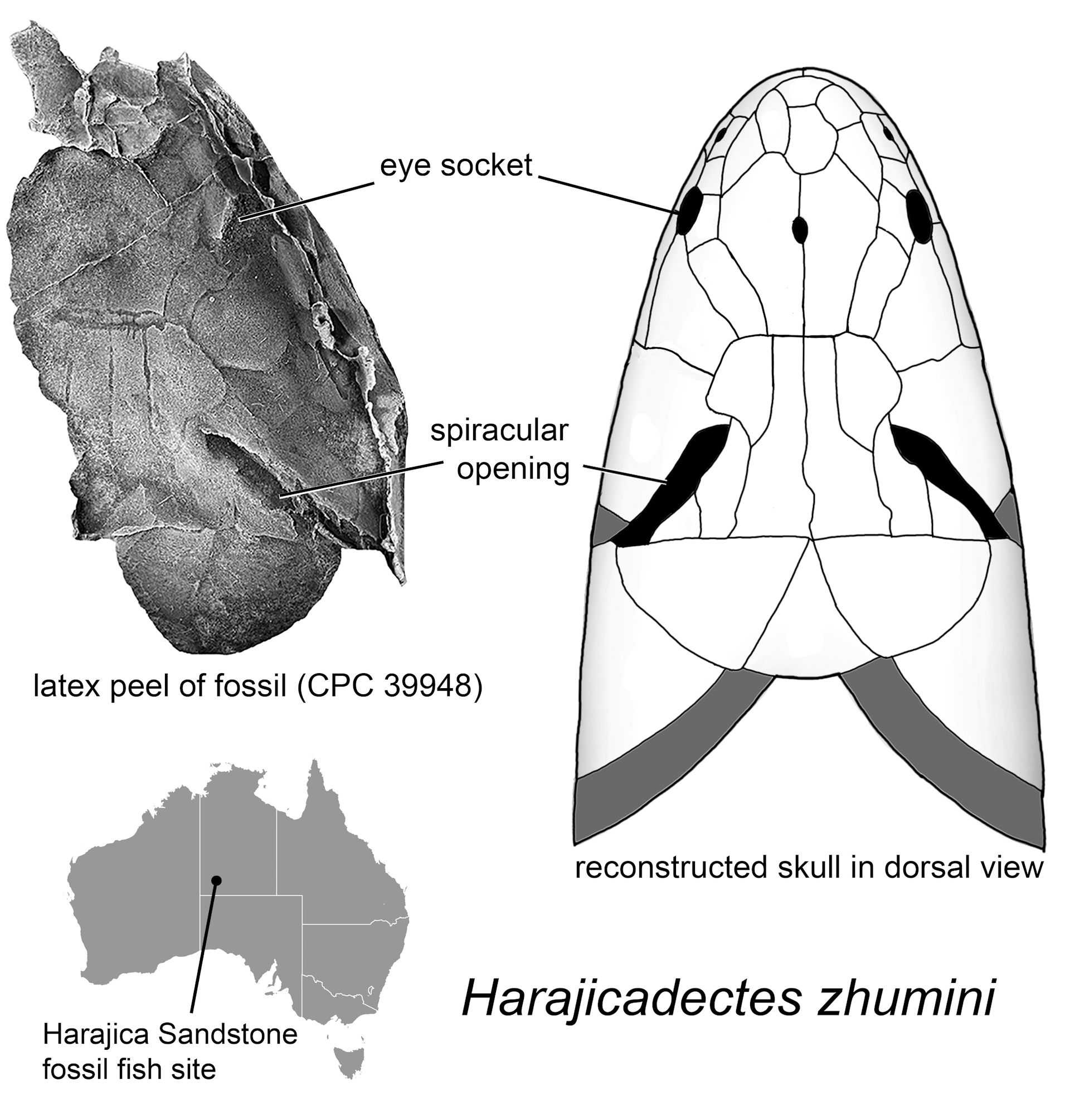
The skull of Harajicadectes seen from above, showing the enormous spiracles.
Credit: Author provided.
That these structures appeared roughly simultaneously in four Devonian lineages provides a fossil “signal” for scientists attempting to reconstruct atmospheric conditions in the distant past.
It could help us uncover the evolution of air breathing in backboned animals.
Brian Choo, Postdoctoral fellow in vertebrate palaeontology, Flinders University; Alice Clement, Research Associate in the College of Science and Engineering, Flinders University, and John Long, Strategic Professor in Palaeontology, Flinders University
This article is republished from The Conversation under a Creative Commons license. Read the original article.
ABSTRACTThe problem with an article like this is that too much detail gives creationists an opportunity to attack the whole thing by focusing on a minor detail, so you can be sure that the few who ever find the courage to read it will be scrutinizing it with that in mind, or looking for something to quote (or misquote) out of context. But if you don't include the details, they'll claim there is no evidence, or it is all based on guesses and speculation, or even that the details were left out because they falsified the claims.
Remote Devonian exposures in central Australia have produced significant but highly fragmentary remains of fish-grade tetrapodomorphs. We describe a new tetrapodomorph from the Middle–Late Devonian (Givetian–Frasnian) Harajica Sandstone Member of the Amadeus Basin, Northern Territory, which is represented by several nearly complete skulls along with much of the body and postcranial skeleton. The new form has a posteriorly broad postparietal shield, broad, triangular extratemporal bones, and a lanceolate parasphenoid. The spiracular openings are particularly large, a character also recorded in elpistostegalians and Gogonasus, demonstrating that these structures, suggestive of spiracular surface air-breathing, appeared independently in widely differing nodes of the stem-tetrapod radiation. A phylogenetic analysis resolves the new form within a cluster of osteolepidid-grade taxa, either as part of a polytomy or as the most basally-branching representative of a clade containing ‘osteolepidids,’ canowindrids, and megalichthyids.
INTRODUCTION
Clade Tetrapodomorpha comprises the limbed tetrapods and their closest fish relatives, whose earliest record is from the Pragian of China (Lu et al., 2012). The group diversified greatly in both marine and freshwater habitats during the Middle-to-Late Devonian while giving rise to several distinct lineages, including the earliest limbed tetrapods (Ahlberg, 2018). Whereas the tetrapods flourished after the Devonian, limbless fish-grade tetrapodomorphs underwent a marked reduction in diversity during the Carboniferous, with only a handful of representatives persisting into the early Permian before vanishing from the fossil record (Romano et al., 2014). Australia is renowned for its excellent fossil record of Devonian tetrapodomorph fishes, with most discoveries in non-marine Givetian–Famennian sites located in the southeast of the continent (Ahlberg & Johanson, 1997; Johanson & Ahlberg, 1997.1, 1998; Long, 1985a, 1985.1b, 1987; Thomson, 1973; Young et al., 2013, 2019). The oldest Australian tetrapodomorph occurrence is in the Emsian–?Eifelian Hatchery Creek Group of Burrinjuck, New South Wales, the material initially provisionally referred to the European genus Gyroptychius by Young and Gorter (1981), but more recently recognized to contain several taxa including a form close to Kenichthys from the Emsian of China (Hunt & Young, 2012.1; Young & Lu, 2020). Outside of this region, the spectacular Devonian reef deposits of the Kimberley region of north-western Western Australia have produced the exceptionally preserved marine vertebrate assemblage of the Gogo Formation (Long & Trinajstic, 2010), which includes Gogonasus Long 1985a, one of the best preserved of all tetrapodomorph fishes (Holland & Long, 2009; Long et al., 1997.2, 2006). In contrast to these exceptional records, fossil collections from the extensive Devonian exposures of central Australia remain poorly sampled yet have already provided significant insights into the early evolution and distribution of tetrapodomorphs in Eastern Gondwana. The fossil record of this region includes the potentially oldest remains referrable to the Rhizodontida, based on fragmentary material from the Early–Middle Devonian Wuttagoonaspis fossil fish assemblage of the Georgina Basin of the Northern Territory and western Queensland (Young & Goujet, 2003). Elsewhere in East Gondwana, the rhizodontid Aztecia occurs in the Middle–Late Devonian of the Transantarctic Mountains (Johanson & Ahlberg, 2001; Young et al., 1992), and the most basally-branching rhizodontid known from reasonably complete remains is Gooloogongia loomesi from the late Frasnian of Canowindra, New South Wales (Johanson & Ahlberg, 1998). Other central Australian discoveries include several forms placed within the ‘Osteolepididae,’ a group now generally considered to represent a paraphyletic assemblage of basally-branching tetrapodomorphs (Ahlberg & Johanson, 1998.1). Muranjilepis winterensis is based on parietal and post-parietal shields from Mount Winter, in the Northern Territory (Young & Lu, 2020; Young & Schultze, 2005). Isolated cosmoid scales with posterior serrations from an unidentified ‘osteolepidid’ occur in the Cravens Peak Beds of the Toomba Range, western Queensland (Young & Schultze, 2005). These sites are dated as Emsian–Eifelian, and thus are also of comparable age to some of the earliest known tetrapodomorph fossils from Yunnan, China, namely Kenichthys campbelli (Chang & Zhu, 1993; Zhu & Ahlberg, 2004) from the Emsian Chuandong Formation, and the oldest and basally-branching representative of the clade, Tungsenia paradoxa from the Pragian Posongchong Formation (Lu et al., 2012). Another highly fragmentary record was a tetrapodomorph first mentioned by Young (1985.2) as a “rhipidistian crossopterygian,” based on fragmentary material collected in 1973 from a fossil fish assemblage at “locality 6” (Fig. 1) in the Harajica Sandstone Member of the Parke Siltstone, part of the non-marine Pertnjara Group of the Amadeus Basin in the Northern Territory (Jones, 1972, 1991). The material comprised a partial maxillary or dentary, an isolated mandibular element, and a denticulate plate presumed to come from the mouth cavity. The Harajica fossil fish assemblage is dominated by the widely distributed antiarch Bothriolepis sp. (Young, 1985.2), associated with the phyllolepid placoderm Placolepis harajica (Young, 1988, 2005.1), two types of acanthodian fin-spines (Young, 1985.2), and the dipnoan Harajicadipterus youngi (Clement, 2009.1).Additional specimens of the Harajica tetrapodomorph were recovered in collecting expeditions in 1991 and 2016 (Figs. 2–6), and this paper describes and analyses this new taxon. The new form is the only tetrapodomorph fish known from central Australia that is represented by articulated fossil material. FIGURE 1. Locality maps for the fossil material described in this work. A, the Amadeus Basin of central Australia. All fossils were collected from locality 6 (redrawn from figure 1 in Young, 1985.2). B, geological map of western MacDonnell Ranges, NT, showing locality 6 about 2 km southwest of Stokes Pass (redrawn from figure 6 in Young, 1985.2). Abbreviations: Pzk, Paleozoic.
FIGURE 1. Locality maps for the fossil material described in this work. A, the Amadeus Basin of central Australia. All fossils were collected from locality 6 (redrawn from figure 1 in Young, 1985.2). B, geological map of western MacDonnell Ranges, NT, showing locality 6 about 2 km southwest of Stokes Pass (redrawn from figure 6 in Young, 1985.2). Abbreviations: Pzk, Paleozoic. FIGURE 2. Harajicadectes zhumini from the Harajica Sandstone Member (Givetian–Frasnian), Northern Territory, Australia, Holotype NTM P6410. A, photographed as a natural mold in situ as it was discovered in 2016; B, as a whitened latex peel; and C, interpretative drawing.
FIGURE 2. Harajicadectes zhumini from the Harajica Sandstone Member (Givetian–Frasnian), Northern Territory, Australia, Holotype NTM P6410. A, photographed as a natural mold in situ as it was discovered in 2016; B, as a whitened latex peel; and C, interpretative drawing. FIGURE 3. Harajicadectes zhumini, skulls in dorsal view, all whitened latex peels of the original fossil molds. A, photographs and B, drawings of NTM P6410 (holotype); C–D, CPC 39948 (paratype); E–F, NMVP 228722; G–H, AMF 99704.
FIGURE 3. Harajicadectes zhumini, skulls in dorsal view, all whitened latex peels of the original fossil molds. A, photographs and B, drawings of NTM P6410 (holotype); C–D, CPC 39948 (paratype); E–F, NMVP 228722; G–H, AMF 99704. FIGURE 4. Harajicadectes zhumini, snout, jaws, and palate. A, photograph and B, drawing of a fragment of the snout in left dorsolateral view, CPC 39949B. C, photograph and D, drawing of isolated mandible, NMVP 253964. E, left maxilla in mesial view, NMV P253965B. F, partial maxilla and lingual surface of the mandible, NMV P253965A. G, photograph, H, drawing, and I, attempted reconstruction of the palate, CPC 33949A.
FIGURE 4. Harajicadectes zhumini, snout, jaws, and palate. A, photograph and B, drawing of a fragment of the snout in left dorsolateral view, CPC 39949B. C, photograph and D, drawing of isolated mandible, NMVP 253964. E, left maxilla in mesial view, NMV P253965B. F, partial maxilla and lingual surface of the mandible, NMV P253965A. G, photograph, H, drawing, and I, attempted reconstruction of the palate, CPC 33949A. FIGURE 5. Harajicadectes zhumini, body and fins. A, photograph and B, drawing of body scales, opercular-gular elements and pectoral girdle, NMVP 229368. C, squamation and D, lepidotrichia from the rear of the body, immediately anterior to the base of the caudal fin, CPC 39953. E–F, squamation from the anterior of the body with partial head, operculum, and pectoral fin, CPC 39952. G–H, mesial view of mid-body squamation with detached pectoral fin and basal scute, CPC 39954.GEOLOGICAL SETTING
FIGURE 5. Harajicadectes zhumini, body and fins. A, photograph and B, drawing of body scales, opercular-gular elements and pectoral girdle, NMVP 229368. C, squamation and D, lepidotrichia from the rear of the body, immediately anterior to the base of the caudal fin, CPC 39953. E–F, squamation from the anterior of the body with partial head, operculum, and pectoral fin, CPC 39952. G–H, mesial view of mid-body squamation with detached pectoral fin and basal scute, CPC 39954.GEOLOGICAL SETTING FIGURE 6. Harajicadectes zhumini. A, scales adjacent to the 2nd dorsal fin, CPC 39953. B, scales from the anterior half of the body, CPC 39952.
FIGURE 6. Harajicadectes zhumini. A, scales adjacent to the 2nd dorsal fin, CPC 39953. B, scales from the anterior half of the body, CPC 39952.
The Harajica Sandstone Member of the Parke Siltstone (Fig. 1) was described in detail by Jones (1972). Young (1985.2) reported a measured section with the fish remains mainly restricted to a 15 m zone 75–90 m above the underlying Deering Siltstone, the basally-branching member of the Pertnjara Group. Young (1985.2) initially estimated a Givetian–Frasnian age for the Harajica fish assemblage. The underlying Deering Siltstone produced a single fish impression with coarse tubercular ornament from a nearby locality (Dare Plain; locality 2 of Young, 1985.2). A better specimen (Young, 1988) from a similar horizon near the base of the Pertnjara Group at Mount Winter is a spine referable to the placoderm Wuttagoonaspis. The Wuttagoonaspis assemblage is widely distributed across the Early–Middle Devonian of central Australia (Young & Goujet, 2003). In the western part of the Amadeus Basin it is known from the base of the Pertnjara Group and upper part of the Mereenie Sandstone (Young & Schultze, 2005; Young & Turner, 2000). Young (1991.1) noted that a Geminospora lemurata spore assemblage, previously reported from Dare Plain, elsewhere ranges down into the Eifelian, which was consistent with a Givetian age for the Amadeus Basin occurrence. Taking into account other microvertebrate evidence, Young and Turner (2000: fig. 4) correlated the Harajica fauna with the Antarctic Aztec fish assemblage (Macrovertebrate zone 6d), of probable Givetian age.
Brian Choo, Timothy Holland, Alice M. Clement, Benedict King, Tom Challands, Gavin Young & John A. Long (2024)
A new stem-tetrapod fish from the Middle–Late Devonian of central Australia,
Journal of Vertebrate Paleontology, DOI: 10.1080/02724634.2023.2285000
Copyright: © 2024 The authors.
Published by Informa UK Limited. Open access
Reprinted under a Creative Commons Attribution 4.0 International license (CC BY 4.0)






What you can almost guarantee is that they will never follow any links to the original science paper and if they do, they won't understand what they're reading or the reason scientists never express certainty but always allow for error and uncertainty. A creationist’s thoughts will always be, "What reasons can I find to dismiss this?", "Why is this wrong?" or "How can I misrepresent this information?", and never, "What are the scientists claiming and have they established their claims with a reasonable degree of certainty, and should I be changing my mind in view of this new information?"
And what they will never admit is that scientists have discovered a 380-million-year-old fossil of a fish that was related to the first terrestrial tetrapods, no matter how compelling the evidence they have presented, because admitting they could be wrong will bring their entire edifice crashing down and leave them nothing to believe in, no excuses for feeling smugly superior to those clever-dickie scientists, and feeling less important than they think they should be.
What Makes You So Special? From The Big Bang To You

How did you come to be here, now? This books takes you from the Big Bang to the evolution of modern humans and the history of human cultures, showing that science is an adventure of discovery and a source of limitless wonder, giving us richer and more rewarding appreciation of the phenomenal privilege of merely being alive and able to begin to understand it all.
Available in Hardcover, Paperback or ebook for Kindle
Ten Reasons To Lose Faith: And Why You Are Better Off Without It

This book explains why faith is a fallacy and serves no useful purpose other than providing an excuse for pretending to know things that are unknown. It also explains how losing faith liberates former sufferers from fear, delusion and the control of others, freeing them to see the world in a different light, to recognise the injustices that religions cause and to accept people for who they are, not which group they happened to be born in. A society based on atheist, Humanist principles would be a less divided, more inclusive, more peaceful society and one more appreciative of the one opportunity that life gives us to enjoy and wonder at the world we live in.
Available in Hardcover, Paperback or ebook for Kindle

No comments:
Post a Comment
Obscene, threatening or obnoxious messages, preaching, abuse and spam will be removed, as will anything by known Internet trolls and stalkers, by known sock-puppet accounts and anything not connected with the post,
A claim made without evidence can be dismissed without evidence. Remember: your opinion is not an established fact unless corroborated.