
Although the horns of cattle, gazelles and goats look very different to the antlers of deer and the ossicones on the head of a giraffe, and indeed, they are constructed differently, the cells they develop from in the embryo are the cells of the 'neural crest' that also develop into the face rather than the rest of the cranium. That and the fact that the underlying genetic control of their growth is sufficiently similar, provides compelling evidence that they share a common origin from which they, and the orders of which they are typical, have diverged.
This is the conclusion of two researchers at the American Museum of Natural History and Baruch College and the CUNY Graduate Center, who have just published their findings, open access in the journal, Communications Biology. It is also explained in an American Museum of Natural History press release:
From the small ossicones on a giraffe to the gigantic antlers of a male moose—which can grow as wide as a car—the headgear of ruminant hooved mammals is extremely diverse, and new research suggests that despite the physical differences, fundamental aspects of these bony adaptations likely evolved from a common ancestor. This finding is published today in the journal Communications Biology by researchers from the American Museum of Natural History and Baruch College and the CUNY Graduate Center.
There are about 170 modern ruminant hooved mammal species with headgear, and many more in the fossil record. The headgear we see today comes in four types—antlers, horns, ossicones, and pronghorns—and they are used in a variety of ways, including for defense, recognition of other members of the species, and mating. Until recently, scientists were unsure if these various bony headgear evolved independently in each ruminant group or from a shared common ancestor.Horns and antlers are incredibly diverse structures, and scientists have long debated their evolutionary origins. This genomic research not only gets us closer to solving an evolutionary mystery, but also helps us better understand how bone forms in all mammals.
Assistant Professor Zachary T. Calamari, Lead author
Department of Natural Sciences
Baruch College, City University of New York, NY, USA.
As a comparative biology Ph.D. student in the Museum’s Richard Gilder Graduate School, Calamari began investigating this question using genomic and computer-based 3D shape analysis. Working with the Museum’s Frick Curator of Fossil Mammals John Flynn, Calamari focused on sequencing transcriptomes, the genes expressed in a tissue at a specific time, for headgear. Their research supports the idea that all of the ruminant headgear forms evolved from a common ancestor as paired bony outgrowths from the animals’ “forehead,” the area near the frontal bones of the skull.
By comparing their newly sequenced cattle horn transcriptome to deer antler and pig skin transcriptomes, Calamari and Flynn confirmed for the first time with transcriptomes that family-specific differences in headgear likely evolved as elaborations on a general bony structure inherited from a common ancestor.Our results provide more evidence that horns form from the cranial neural crest, an embryonic cell layer that forms the face, rather than from the cells that form the bones on the sides and back of the head, It is striking that these are the same cells that form antlers. And the distinctive patterns of gene expression in cattle horns and deer antlers, relative to other bone and skin tissue “controls,” provide compelling evidence of shared origin of fundamental aspects of these spectacular bony structures in an ancient ancestor.
John J. Flynn, co-author
Richard Gilder Graduate School
American Museum of Natural History, New York, NY, USA.In addition to gene expression patterns that support a single origin of horns and antlers, our results also show the regulation of gene expression patterns in these structures may differ from other bones. These results help us understand the evolutionary history of horns and antlers and could suggest that differences in other ruminant cranial appendages, like ossicones and pronghorns, are also elaborations on a shared ancestral cranial appendage.
Assistant Professor Zachary T. Calamari.
AbstractSo, yet more evidence of common origins and descent with modification, and still no sign that the impending abandonment of the Theory of Evolution, of which this evidence is consistent, by professional biologists who will soon adopt creationism with its supernatural magic and unproven entities, as the creation cult leaders have been fooling their dupes into thinking will happen any day now, real soon, for the last half a century.
Horns, antlers, and other bony cranial appendages of even-toed hoofed mammals (ruminant artiodactyls) challenge traditional morphological homology assessments. Cranial appendages all share a permanent bone portion with family-specific integument coverings, but homology determination depends on whether the integument covering is an essential component or a secondary elaboration of each structure. To enhance morphological homology assessments, we tested whether juvenile cattle horn bud transcriptomes share homologous gene expression patterns with deer antlers relative to pig outgroup tissues, treating the integument covering as a secondary elaboration. We uncovered differentially expressed genes that support horn and antler homology, potentially distinguish them from non-cranial-appendage bone and other tissues, and highlight the importance of phylogenetic outgroups in homology assessments. Furthermore, we found differentially expressed genes that could support a shared cranial neural crest origin for horns and antlers and expression patterns that refine our understanding of the timing of horn and antler differentiation.
Introduction
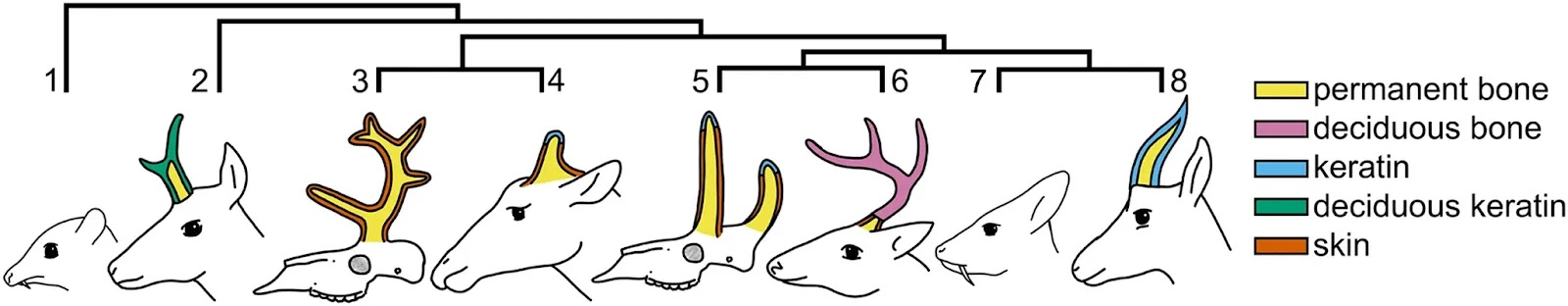
Homology is difficult to establish for the horns, antlers, ossicones, and pronghorns (cranial appendages) of extant ruminants (Artiodactyla, Ruminantia), the clade of even-toed hoofed mammals (cattle, antelopes, deer, and relatives). Biologists use different names for the cranial appendages of each living ruminant family because their tissue composition and growth are distinct (Fig. 1). Nevertheless, all ruminant cranial appendages include permanent bony outgrowths of the frontal and parietal bones with family-specific integument coverings1,2,3. Although some researchers use these morphological and developmental differences to infer independent origins for the distinct types of cranial appendages1, both phylogenetic optimizations based on alternative concepts of cranial appendages and genomic data instead may support a single origin and deep homology of cranial appendages in ruminants4,5,6. Both interpretations are plausible, yet their implications for understanding ruminant craniofacial biology and morphological diversification are profound. In this study, we focus on the horns of Bovidae (e.g., antelopes, goats, cattle) and the antlers of Cervidae (e.g., moose, white-tailed deer, reindeer) to address the question of homology with high-throughput RNA sequencing analyses. These families have the greatest species diversity of living ruminants and include common livestock and game species, which provides better access to their cranial appendage tissues than is possible with the endangered Giraffidae (giraffes and okapi with ossicone appendages) and wild Antilocapridae (pronghorn “antelopes” with pronghorn appendages).
Bovid horn growth appears to begin in a specialized region of the dermis and subcutaneous loose connective tissue (SLCT) overlying the frontal bone of the skull, often called the os cornu1. Although inexact use of the term os cornu has confounded efforts to determine whether it is a separate ossification center1,3, recent examinations show substantial increases in nerve fibers and other morphological changes in developing horn bud dermis relative to other skin overlying the frontal bone7. The tissues clearly promote horn growth through intramembranous ossification1; however, they may not convert epidermis into the keratin sheath, suggesting horn bud and keratin sheath tissues receive these fates separately in utero1,7,8,9. Although the horn bud is distinct from frontal skin in fetal cattle by the second month of gestation, ossification of the bone core only occurs after birth7.Fig. 1: Cranial appendage tissue composition and phylogenetic relationships of living and extinct ruminants.Living and entirely extinct (†) ruminant families (even-toed hoofed mammals; Artiodactyla, Ruminantia), showing cranial appendage types and tissue composition. 1: Tragulidae, 2: Antilocapridae (pronghorns), 3: Climacoceratidae†, 4: Giraffidae (ossicones), 5: Palaeomerycidae†, 6: Cervidae (antlers), 7: Moschidae, 8: Bovidae (horns). Tragulidae and Moschidae lack cranial appendages. Phylogeny based on Hassanin et al.17.
In contrast, the periosteum covering the frontal bone, not the more superficial SLCT and dermis, initiates antler growth1,10,11. During early growth of the pedicle, the permanent bone base of an antler, ossification is intramembranous; later pedicle growth and subsequent antler growth from the pedicle are both endochondral1,11. As with horns, the tissues that give rise to the antler receive this fate in utero, differentiating from surrounding tissues by the second month of gestation11. Unlike the early postnatal onset of horn growth in bovids, pedicle and antler growth in most cervids does not begin until puberty11.
Debate over the origins of cranial appendages involves two competing concepts of cranial appendages as anatomical structures. The first concept includes any family-specific bone or integument covering the permanent bony portion as an essential part of the structure. This “essential integument” concept emphasizes several differences between cranial appendage types to argue that each must have originated independently, such as the tissues that appear to initiate their growth, the type of ossification characterizing their main growth, and the timing of onset of growth. Under the essential integument concept, the differences in horn and antler development and integument covering become definitive indicators of non-homology—a horn cannot have shared origins with an antler, because no known fossil or living cervid has a keratin sheath on its cranial appendages and no known bovid has ever had a deciduous bone shed and regrown annually from its cranial appendage. This concept requires some ruminant families to have evolved cranial appendages multiple times. For example, the extinct merycodontine antilocaprids likely had skin covering their cranial appendages, while their extant relative, Antilocapra americana, has deciduous keratin sheaths covering its cranial appendages1,12. The essential integument concept treats these as two separate originations, solely because of the difference in their integument covering. The presence of a bony core in all antilocaprids, however, indicates that the group ancestrally possessed a bony cranial appendage through simple character optimization, and the evolution of a merycodontine cranial appendage into one like that of A. americana could have occurred simply through changes to the integument covering this bony core12.
Contrasting the essential integument concept is the hypothesis that each cranial appendage evolved by modification of a shared rudimentary ancestral cranial appendage. Under this concept, the permanent bone portion of each cranial appendage is a “homologous core” structure and was present as some rudimentary outgrowth in the common ancestor of all ruminants. The differences between each cranial appendage type, such as the permanent keratin sheath of a horn or the deciduous skin-covered bony antler, thus are independently evolved elaborations upon this common ancestral bony base13. The potentially separate fate determination of the horn bud and keratin sheath during embryonic development7,8,9 could be evidence that the keratin sheath evolved independently from the bony core. The similar position on the skull and initial stages of intramembranous ossification of both horns and antlers (as well as ossicones and pronghorns of other ruminants not included in this study) also would reflect a shared origin of cranial appendages from an ancestor with a rudimentary, intramembranously ossified bony frontal outgrowth. Recently discovered stem fossils from cranial appendage-bearing families evince rudimentary versions of their respective cranial appendages14,15. They demonstrate the potential for fossil discoveries to provide morphological support for the rudimentary ancestral cranial appendage of all ruminants proposed by this hypothesis. In the current paucity of such fossil evidence, shared and derived genetic mechanisms that initiate and direct cranial appendage growth in living ruminants can provide the best evidence for the homologous core concept.
Regardless of which cranial appendage concept applies, homology is fundamentally a question of phylogenetic optimization of traits, be they transcriptomic, genomic, or anatomical. Phylogenetic analyses of molecular data provide well-supported hypotheses of ruminant family interrelationships (Fig. 1) that prompt re-examination of cranial appendage homology3. Notably, these studies find cranial appendage-lacking Moschidae (musk deer) nested within cranial appendage-bearing ruminants as the closest relative to horn-bearing Bovidae16,17,18 rather than outside the living families with cranial appendages. Although both positions support a most parsimonious ancestral origin for cranial appendages in ruminants, a Moschidae + Bovidae clade introduces the possibility of more complicated scenarios of gains and losses. Likewise, molecular phylogenetic analyses place antler-lacking water deer Hydropotes inermis well within the antlered cervids as the closest relative to the antler-bearing roe deer genus Capreolus, rather than as the nearest outgroup to all antler-bearing cervids17,19, rejecting the possibility that this species supports an antlerless origin for Cervidae. Under the homologous core concept, the most parsimonious interpretation of antlerless Hydropotes would be a secondary loss of antlers regardless of where it falls in the cervid phylogeny, because the bony antler pedicle would be inherited from the common ancestor of all cranial appendage-bearing ruminants. Overall, these current phylogenetic positions of Moschidae and Hydropotes require only a single origin of cranial appendages within ruminants and separate losses in these two groups. Separate origins for each cranial appendage type, inclusive of its elaborations, based on the a priori adoption of the essential integument concept, instead requiring four or more gains or losses of cranial appendages across ruminants.
Just as the addition of molecular evidence has altered our understanding of ruminant evolutionary relationships, it also can enhance our understanding of cranial appendage homology. If cranial appendages first arose as a rudimentary bony structure in the ruminant common ancestor, shared biological processes may initiate and sustain their growth in all extant ruminants. These shared processes should be evident through the genes expressed during development1,3,20,21,22. High-throughput sequencing studies of horns or antlers, however, have focused on these structures within a single species or type of appendage23,24. The few studies of gene expression in cranial appendages that have compared different cranial appendage types provide valuable insights into genes that may support homology but lack the outgroup comparisons necessary for a rigorous, phylogeny-constrained test of homology4,5,6.
Additionally, the timing and rate of development of structures can evolve, and these changes are often responsible for evolving novel morphology25. For example, changing when tooth formation begins may result in teeth appearing to form from different layers of oral or dental epithelia, even though the teeth themselves are homologous structures26. A similar heterochronic shift may be responsible for developmental differences in timing and tissue layer observed between cranial appendages. Antler growth and regeneration are driven by stem cells in the periosteum11; although stem cells have not yet been identified in horn growth, formation of the os cornu, horn core, and keratin sheath likely involve some aspect of mesenchymal cell differentiation. In particular, both horn and antler precursors likely form from cranial neural crest cells6,11,23,27, although direct experimental confirmation is lacking. Regardless, developmental differences or similarities between two groups are not sufficient by themselves to reject or establish homology; they also must be considered within the context of the ancestral conditions and subsequent modifications reconstructed through phylogenetic outgroup comparisons.
In this study, we assessed the homology of cranial appendages under the homologous core concept, testing the hypothesis that the horn buds of juvenile cattle are homologous to antler pedicle tissues. We compared genes expressed by a developmental sequence of juvenile cattle horn buds to those expressed by deer antler pedicles, antler tips, and isolated antler tip tissues to explore heterochronic changes in expression timing that may support homology. We included a baseline outgroup comparison to pig skin, cartilage, and bone to assess homology. We predicted that the proportion of homologously expressed genes shared between horn buds and antler tissues would decrease over development, reflecting an evolutionarily conserved early trajectory for the permanent bone base that only differentiates later in development to yield family-specific features. Horn buds are essentially skin that can induce additional bone growth, so we expected a significant overlap with genes differentially expressed (DE) in the skin. Focusing on genes and pathways that differentiate cranial appendages from these within-species skin controls, we identified multiple genes with homologous expression patterns between cattle horns and deer antlers. Our results support the homology of horns and antlers through gene expression and highlight the need to include a phylogenetic outgroup in homology assessments using genomics data. Our results also may support cranial neural crest derivation of horns and suggest regulatory genes of interest for future analyses of cranial appendage evolution.
Calamari, Z.T., Flynn, J.J.
Gene expression supports a single origin of horns and antlers in hoofed mammals. Commun Biol 7, 509 (2024). https://doi.org/10.1038/s42003-024-06134-4
Copyright: © 2024 The authors.
Published by Springer Nature Ltd. Open access.
Reprinted under a Creative Commons Attribution 4.0 International license (CC BY 4.0)
Meanwhile, the mountain of evidence for evolution continues to grow...
What Makes You So Special? From The Big Bang To You

Ten Reasons To Lose Faith: And Why You Are Better Off Without It


No comments:
Post a Comment
Obscene, threatening or obnoxious messages, preaching, abuse and spam will be removed, as will anything by known Internet trolls and stalkers, by known sock-puppet accounts and anything not connected with the post,
A claim made without evidence can be dismissed without evidence. Remember: your opinion is not an established fact unless corroborated.