

Their findings are published, open access, in the journal Nature and are described in a Penn State, Eberly College news release:
Technical details are given in the team's abstract and introduction to their open access paper in Nature:New end-to-end sequences uncover enormous variation on the Y chromosome, informing human evolution and disease as well as conservation genetics of endangered apes
Newly generated, complete “end-to-end” reference genomes for the sex chromosomes of five great ape species and one lesser ape species — produced by an international collaborative team led by researchers at Penn State, the National Human Genome Research Institute and the University of Washington — highlight extremely rapid changes on the male-specific Y chromosome among ape species. These findings shed light on the evolution of sex chromosomes and inform understanding of diseases related to genes on these chromosomes in both apes and humans.
The Y chromosome is important for human fertility, and the X chromosome harbors genes critical for reproduction, cognition and immunity. Our study opens doors for many future investigations of sex chromosomes, how they evolved and diseases associated with them. The living non-human great ape species we studied are all endangered. The availability of their complete sex chromosome sequences will facilitate studies of their sex-specific dispersal in the wild and of their genes important for reproduction and fertility.
Professor Kateryna Makova, co-first author
Verne M. Willaman Chair of Life Sciences
Director of the Center for Medical Genomics
Penn State University, PA, USA.
Such reference genomes act as a representative example that are useful for future studies of these species. The team found that, compared to the X chromosome, the Y chromosome varies greatly across ape species and harbors many species-specific sequences. However, it is still subject to purifying natural selection — an evolutionary force that protects its genetic information by removing harmful mutations.
The new study appeared today (May 29) in the journal Nature.
The team produced complete sex chromosome sequences for five species of great apes — chimpanzee, bonobo, gorilla, Bornean orangutan and Sumatran orangutan, which comprise most great ape species living today — as well as a lesser ape, siamang. They generated sequences for one individual of each species. The resulting reference genomes act as a map of genes and other chromosomal regions, which can help researchers sequence and assemble the genomes of other individuals of that species. Previous sex chromosome sequences for these species were incomplete or — for the Bornean orangutan and siamang — did not exist.Researchers sequenced the human genome in 2001, but it wasn’t actually complete. The technology available at the time meant that certain gaps weren’t filled in until a renewed effort led by the Telomere-to-Telomere, or T2T, Consortium in 2022-23. We leveraged the experimental and computational methods developed by the Human T2T Consortium to determine the complete sequences for the sex chromosomes of our closest living relatives—great apes.
Professor Kateryna Makova.
High variability on the Y chromosomeThe Y chromosome has been challenging to sequence because it contains many repetitive regions, and, because traditional short-read sequencing technology decodes sequences in short bursts, it is difficult to put the resulting segments in the correct order. T2T methods use long-read sequencing technologies that overcome this challenge. Combined with advances in computational analysis, on which we collaborated with Adam Phillippy’s group at the NHGRI, this allowed us to completely resolve repetitive regions that were previously difficult to sequence and assemble. By comparing the X and Y chromosomes to each other and among species, including to the previously generated human T2T sequences of the X and the Y, we learned many new things about their evolution.
Dr Karol Pál, co-first author.
Penn State University, PA, USA.As a result, across the six ape species, the research team found that the Y chromosome was much more variable than the X over a variety of characteristics, including size. Among the studied apes, the X chromosome ranges in size from 154 million letters of the ACTG alphabet — representing the nucleotides that make up DNA — in chimpanzee and human to 178 million letters in gorilla. In contrast, the Y chromosome ranges from 30 million DNA letters in siamang to 68 million letters in Sumatran orangutan.Sex chromosomes started like any other chromosome pair, but the Y has been unique in accumulating many deletions, other mutations and repetitive elements because it does not exchange genetic information with other chromosomes over most of its length.
Professor Kateryna Makova.
The amount of DNA sequence shared between species was also more variable on the Y. For example, about 98% of the X chromosome aligns between human and chimpanzee, but only about a third of the Y aligns between them. The researchers found that this is in part because the Y chromosome is more likely to be rearranged or have portions of its genetic material duplicated.
Additionally, the percentage of the chromosome occupied by sequences that are repeated is highly variable on the Y. Whereas, depending on the species, 62% to 66% of the X chromosomes are occupied by repetitive elements, 71% to 85% of the Y chromosomes are occupied by them. These percentages are higher on both the X and the Y than in other chromosomes in the human genome.
How the Y has survivedThe researchers found that many genes on the Y chromosome seem to use two strategies to survive. The first takes advantage of genetic redundancy — the presence of multiple copies of the same gene on a chromosome — so that intact copies of the gene can compensate for copies that might acquire mutations. The team quantified this genetic redundancy by completing the landscape of multi-copy gene families on ape sex chromosomes for the first time.We found the ape Y to be shrinking, accumulating many mutations and repeats, and losing genes. So why hasn’t the Y chromosome disappeared, as some previous hypotheses suggested? In collaboration with Sergei Kosakovsky Pond from Temple University and others, we found that the Y chromosome still has a number of genes evolving under purifying selection — a type of natural selection that keeps gene sequences intact. Many of these genes are important for spermatogenesis. This means that the Y chromosome is unlikely to disappear any time soon.
Professor Kateryna Makova.
What are palindromes in the context of DNA sequences in chromosomes and what is their significance? Palindromes in the context of DNA sequences are segments of DNA that read the same in both directions (5' to 3' and 3' to 5'). Specifically, they are sequences of base pairs that exhibit symmetry such that the sequence on one strand matches the complementary sequence on the opposite strand when read in the reverse direction.The second survival strategy takes advantage of palindromes, where the sequence of letters in the DNA alphabet is followed by the same, but inverted sequence, for example, ACTG-GTCA. When located within a palindrome, genes benefit from the palindrome’s ability to correct mutations.
Structure of DNA Palindromes
In DNA palindromes, the two complementary strands form a sequence such that if one strand has a sequence, the opposite strand will have the complementary sequence read in the reverse direction. For instance, a simple palindromic sequence might be:
5' - GAATTC - 3'
3' - CTTAAG - 5'
When read in the reverse direction, the sequence on the first strand (5' to 3') is complementary to the sequence on the second strand (3' to 5').
Significance of DNA Palindromes
- Restriction Enzymes Recognition Sites: Many restriction enzymes recognize and cut DNA at specific palindromic sequences. These enzymes are crucial tools in molecular biology for DNA cloning, mapping, and analysis. For example, the restriction enzyme EcoRI recognizes the palindromic sequence GAATTC.
- DNA Replication and Repair: Palindromic sequences can form secondary structures, such as hairpins and cruciforms, due to their symmetry. These structures play roles in DNA replication and repair mechanisms. However, they can also cause genomic instability if not properly managed.
- Gene Regulation: Palindromic sequences can be involved in the regulation of gene expression. Some regulatory proteins and transcription factors bind specifically to palindromic DNA sequences to activate or repress transcription.
- Genomic Stability: While palindromic sequences can have regulatory roles, they are also prone to forming secondary structures that can lead to genomic instability, such as deletions, duplications, and translocations. This instability can be a source of genetic diversity but can also lead to diseases if not properly controlled.
- Evolutionary Conservation: Some palindromic sequences are highly conserved across species, indicating their importance in crucial biological functions and evolutionary conservation. These sequences may be involved in essential cellular processes and thus maintained throughout evolution.
In summary, DNA palindromes are important for various biological processes, including enzyme recognition, gene regulation, and maintaining genomic stability. Their symmetrical nature allows them to form unique structures and interact with specific proteins, making them significant in both normal cellular functions and disease states.
We found that the Y chromosome can exchange genetic information with itself between the repeated sequences of the two palindrome arms, which fold so that the inverted sequences align. When two copies of the same gene are located within palindromes, and one copy is hit by a mutation, the mutation can be rescued by the genetic exchange with another copy. This can compensate for the Y’s lack of genetic information exchange with the other chromosomes.
Dr Karol Pál.
The research team obtained the complete sequences of palindromes on ape sex chromosomes also for the first time, as they were previously difficult to sequence and study. They found that palindromes are particularly abundant and long on the ape Y chromosome, yet they are usually only shared among closely related species.
In collaboration with Michael Schatz and his team at Johns Hopkins University, the researchers also studied the sex chromosomes of 129 individual gorillas and chimpanzees to better understand the genetic variation within each species and search for evidence of natural selection and other evolutionary forces acting on them.
We obtained substantial new information from previously studied gorilla and chimpanzee individuals by aligning their sex chromosome sequencing reads to our new reference sequences. While increasing the sample size in the future will be very helpful to improve our ability to detect signatures of different evolutionary forces, this can be ethically and logistically challenging when working with endangered species, so it is critical that we can get the most out of the data we do have.
Assistant Professor Zachary Szpiech, co-author
Penn State University, PA, USA.
The researchers explored a variety of factors that could explain variation on the Y chromosome within gorillas and within chimpanzees, and this analysis revealed additional signatures of purifying selection on the Y. This confirms the role of this type of natural selection on the Y, as was discovered in their previous analyses of genes.
The powerful combination of bioinformatic techniques and evolutionary analyses that we used allows us to better explain the evolutionary processes acting on sex chromosomes in our closest living relatives, great apes. Additionally, the reference genomes we produced will be instrumental for future studies of primate evolution and human diseases.
Assistant Professor Christian Huber, co-author
Penn State University, PA, USA.
In addition to Makova, Pál, Szpiech and Huber, the research team at Penn State includes Kaivan Kamali, computational scientist in the departments of biology and of biochemistry and molecular biology; Troy LaPolice, graduate student in bioinformatics and genomics; Paul Medvedev, professor of computer science and engineering and of biochemistry and molecular biology; Sweetalana, research assistant in the department of biology; Huiqing Zeng, research technologist in biology; Xinru Zhang, graduate student in bioinformatics and genomics; Robert Harris, assistant research professor of biology, now retired; Barbara McGrath, associate research professor of biology, now retired; and Sarah Craig, associate research professor of biology, currently a program officer at the National Institutes of Health. The co-authors also included Penn State alumni Monika Cechova, currently a postdoctoral fellow at the University of California Santa Cruz, and Melissa Wilson, currently an associate professor at Arizona State University.
In addition to Makova, the team was co-led by co-corresponding study authors Adam Phillippy, senior investigator at NHGRI, and Evan Eichler, professor of Genome Sciences at the University of Washington. A full list of authors for this paper is available here.
[My highlights]
Credit: Makova Lab.
AbstractCreationists should make a special note to either ignore or think of a way to misrepresent the following from this paper:
Apes possess two sex chromosomes—the male-specific Y chromosome and the X chromosome, which is present in both males and females. The Y chromosome is crucial for male reproduction, with deletions being linked to infertility1. The X chromosome is vital for reproduction and cognition2. Variation in mating patterns and brain function among apes suggests corresponding differences in their sex chromosomes. However, owing to their repetitive nature and incomplete reference assemblies, ape sex chromosomes have been challenging to study. Here, using the methodology developed for the telomere-to-telomere (T2T) human genome, we produced gapless assemblies of the X and Y chromosomes for five great apes (bonobo (Pan paniscus), chimpanzee (Pan troglodytes), western lowland gorilla (Gorilla gorilla gorilla), Bornean orangutan (Pongo pygmaeus) and Sumatran orangutan (Pongo abelii)) and a lesser ape (the siamang gibbon (Symphalangus syndactylus)), and untangled the intricacies of their evolution. Compared with the X chromosomes, the ape Y chromosomes vary greatly in size and have low alignability and high levels of structural rearrangements—owing to the accumulation of lineage-specific ampliconic regions, palindromes, transposable elements and satellites. Many Y chromosome genes expand in multi-copy families and some evolve under purifying selection. Thus, the Y chromosome exhibits dynamic evolution, whereas the X chromosome is more stable. Mapping short-read sequencing data to these assemblies revealed diversity and selection patterns on sex chromosomes of more than 100 individual great apes. These reference assemblies are expected to inform human evolution and conservation genetics of non-human apes, all of which are endangered species.
Main
Therian X and Y chromosomes are thought to have originated from a pair of autosomes around 170 million years ago3. The X chromosome, which is typically present as two copies in females and one copy in males, has mostly retained the gene content and order from the original autosomal pair4. The Y chromosome, which is typically present as one copy in males, has acquired the sex-determining gene SRY and other male-specific genes and mutations, which were fixed by inversions that prevented recombination between the Y and X chromosomes over most of their lengths5,6. Lacking recombination, the Y chromosome has contracted in size and accumulated deleterious mutations and repetitive elements, leading to differences in size and gene content between the Y and X chromosomes. The recent human T2T (gapless and complete) assembly revealed an X chromosome of around 154 Mb with 796 protein-coding genes7, and a Y chromosome of around 62 Mb with 106 protein-coding genes8. In addition to the pseudoautosomal regions (PARs), where the Y chromosome still recombines with the X chromosome, and ancestral regions, which originated from the original autosomal pair, the human Y chromosome has long ampliconic regions with extensive intrachromosomal homology. Ampliconic regions harbour palindromes—long inverted repeats that undergo gene conversion, which counteracts the accumulation of deleterious mutations9. Similar to the human Y chromosome, the human X chromosome possesses PARs7, ancestral regions and several palindromes10.
Whereas human sex chromosomes have recently been completely sequenced7,8, the sex chromosomes in our closest relatives—non-human apes—remain incompletely characterized. Owing to the haploid nature and high repetitive element content of the Y chromosome, most previous studies have assembled female genomes, omitting the Y chromosome altogether11. Ape Y chromosomes have sometimes been sequenced with targeted methods6,12,13 or via shotgun sequencing of male genomes14,15, but such assemblies are usually fragmented, collapsed and incomplete. Ape X chromosomes have been deciphered to a greater level of contiguity (for example, in refs. 16,17,18), but their assemblies—particularly for long satellite arrays—have remained unfinished, preventing their complete characterization.
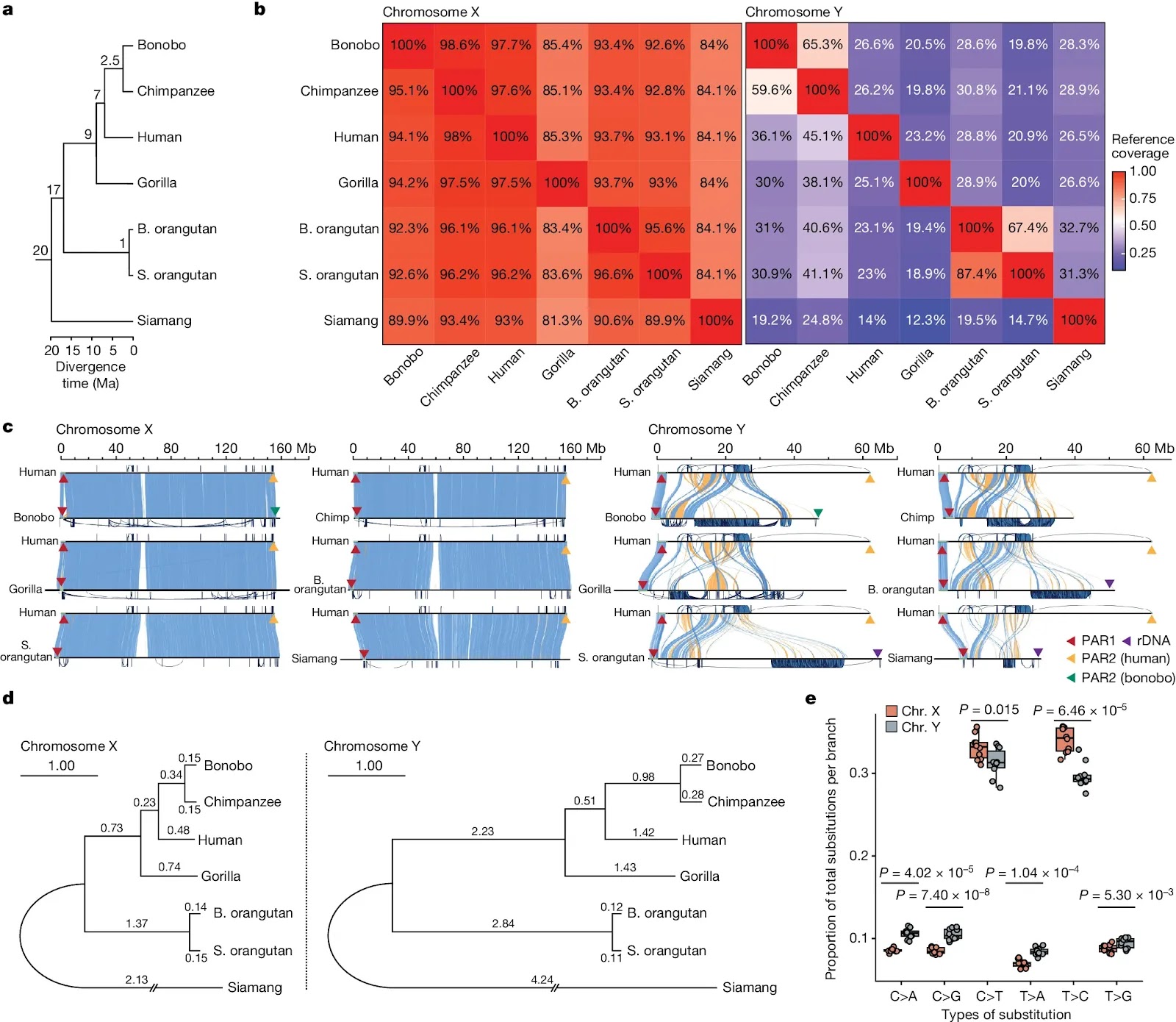
Previous cytogenetic studies demonstrated lineage-specific amplifications and rearrangements leading to large size variations among great ape Y chromosomes (for example, ref. 19). The initial assemblies of the human and chimpanzee Y chromosomes revealed remarkable differences in structure and gene content6,12 despite short divergence time, and an acceleration of substitution rates and gene loss on the Y chromosome was observed in the common ancestor of bonobo and chimpanzee15. The Y chromosome of the common ancestor of great apes probably already possessed ampliconic sequences and multi-copy gene families15, and all ape sex chromosomes share the same evolutionary strata14 while experiencing lineage-specific expansions and loss of ampliconic genes14,15. This progress notwithstanding, the lack of complete ape sex chromosome assemblies has prevented detailed inquiries into the evolution of ampliconic regions, palindromes, segmental duplications, structural variants, satellites, transposable elements and gene copy number. Here, utilizing the experimental and computational methods developed for the T2T assembly of the human genome8,20, we deciphered the complete sequences of sex chromosomes from six ape species and studied their structure and evolution.Fig. 1: Chromosome alignability and divergence. a, The phylogenetic tree of the species in the study (see Supplementary Table 1 for references of divergence times). b, Pairwise alignment coverage of X and Y chromosomes (percentage of reference, as shown on the x axis, covered by the query, as shown on the y axis). c, Alignment of ape sex chromosomes against the human T2T assembly8,20. Blue and yellow bands indicate direct or inverted alignments, respectively. PARs and ribosomal DNA arrays (rDNA) are indicated by triangles (not to scale). Intrachromosomal segmental duplications are drawn outside the axes. The scale bars are aligned to the human chromosome. rDNA, ribosomal DNA. d, Phylogenetic trees of nucleotide sequences on the X and Y chromosomes69. Branch lengths (substitutions per 100 sites) were estimated from multi-species alignment blocks including all seven species. e, A comparison of the proportions of six single-base nucleotide substitution types among total nucleotide substitutions per branch between X and Y (excluding PARs). The distribution of the proportion of each substitution type across 10 phylogenetic branches is shown as a dot plot (all data points are plotted) over the box plot. Box plots show the median as the centre line and the first and third quartiles as bounds; the whiskers extend to the closer of the minimum and maximum value or 1.5 times the interquartile range. The significance of differences in means of substitution proportions between X and Y chromosomes for each substitution type was evaluated with a two-sided t-test on the data from all ten branches (Bonferroni correction for multiple testing was applied).
a, The phylogenetic tree of the species in the study (see Supplementary Table 1 for references of divergence times). b, Pairwise alignment coverage of X and Y chromosomes (percentage of reference, as shown on the x axis, covered by the query, as shown on the y axis). c, Alignment of ape sex chromosomes against the human T2T assembly8,20. Blue and yellow bands indicate direct or inverted alignments, respectively. PARs and ribosomal DNA arrays (rDNA) are indicated by triangles (not to scale). Intrachromosomal segmental duplications are drawn outside the axes. The scale bars are aligned to the human chromosome. rDNA, ribosomal DNA. d, Phylogenetic trees of nucleotide sequences on the X and Y chromosomes69. Branch lengths (substitutions per 100 sites) were estimated from multi-species alignment blocks including all seven species. e, A comparison of the proportions of six single-base nucleotide substitution types among total nucleotide substitutions per branch between X and Y (excluding PARs). The distribution of the proportion of each substitution type across 10 phylogenetic branches is shown as a dot plot (all data points are plotted) over the box plot. Box plots show the median as the centre line and the first and third quartiles as bounds; the whiskers extend to the closer of the minimum and maximum value or 1.5 times the interquartile range. The significance of differences in means of substitution proportions between X and Y chromosomes for each substitution type was evaluated with a two-sided t-test on the data from all ten branches (Bonferroni correction for multiple testing was applied).
Fig. 2: Sequences gained, non-B-DNA, genes, sequence classes, palindromes and intrachromosomal similarity in the assemblies. Tracks for newly generated sequence (black) relative to previous assemblies, non-B-DNA density, gene density (up to 11 genes per 100-kb window), sequence classes (seq. class) and palindromes (black). The X and Y chromosomes are portrayed on different scales. No previous references existed for the Bornean orangutan or siamang, thus the solid black bars for the new sequence tracks. No new sequence was added to the existing T2T human reference in this study and thus the new sequence tracks are empty (white). The gene density tracks are normalized across all species and chromosomes; the non-B-DNA density tracks are calibrated independently for each chromosome; in both cases, darker shades indicate higher density. Self-similarity dot plots using a modified version of Stained Glass70 are shown for the Y chromosomes; satellite arrays are visible as blocks of colour, segmental duplications appear as horizontal lines, and inverted or palindrome repeats are shown as vertical lines.
Tracks for newly generated sequence (black) relative to previous assemblies, non-B-DNA density, gene density (up to 11 genes per 100-kb window), sequence classes (seq. class) and palindromes (black). The X and Y chromosomes are portrayed on different scales. No previous references existed for the Bornean orangutan or siamang, thus the solid black bars for the new sequence tracks. No new sequence was added to the existing T2T human reference in this study and thus the new sequence tracks are empty (white). The gene density tracks are normalized across all species and chromosomes; the non-B-DNA density tracks are calibrated independently for each chromosome; in both cases, darker shades indicate higher density. Self-similarity dot plots using a modified version of Stained Glass70 are shown for the Y chromosomes; satellite arrays are visible as blocks of colour, segmental duplications appear as horizontal lines, and inverted or palindrome repeats are shown as vertical lines.
Fig. 3: Conservation of palindromes and gene density in different sequences classes. a, Palindromes are shown as horizontal lines perpendicular to the chromosomes (painted with sequence classes); palindromes shared among species are connected by coloured lines (different colours are used for unique species combinations, may be dashed when horizontally passing through species without sharing, opacity reduced in regions with dense palindrome sharing). Several gene families that expanded in lineage-specific palindromes on the Y (CDY and RBMY) and that are present in palindromes shared among species on the X chromosome (CENPVL1, FAM156, ETD, HSFX and H2A) are indicated. See Supplementary Tables 36, 37, 38 and 41 for the original data. b, Gene density for different sex chromosome sequence classes. The significance of differences in gene densities was computed using goodness of fit (chi-squared) test with Bonferroni correction for multiple tests. Asterisks indicate significant differences in gene density (P < 0.05). See Supplementary Table 38 for the original data and P values. An interactive version of this plot can be found at https://observablehq.com/d/6e3e88a3e017ec21.
a, Palindromes are shown as horizontal lines perpendicular to the chromosomes (painted with sequence classes); palindromes shared among species are connected by coloured lines (different colours are used for unique species combinations, may be dashed when horizontally passing through species without sharing, opacity reduced in regions with dense palindrome sharing). Several gene families that expanded in lineage-specific palindromes on the Y (CDY and RBMY) and that are present in palindromes shared among species on the X chromosome (CENPVL1, FAM156, ETD, HSFX and H2A) are indicated. See Supplementary Tables 36, 37, 38 and 41 for the original data. b, Gene density for different sex chromosome sequence classes. The significance of differences in gene densities was computed using goodness of fit (chi-squared) test with Bonferroni correction for multiple tests. Asterisks indicate significant differences in gene density (P < 0.05). See Supplementary Table 38 for the original data and P values. An interactive version of this plot can be found at https://observablehq.com/d/6e3e88a3e017ec21.
Fig. 4: Repeats on ape sex chromosomes. a, Repeat annotations across each ape sex chromosome are depicted as a percentage of total nucleotides. Previously uncharacterized human repeats derived from the CHM13 genome analyses are shown in teal. Newly defined satellites (Methods) are depicted in light orange. b, The amount of DNA on each sex chromosome comprising canonical satellites, with each satellite represented by a different colour. LINE, long interspersed nuclear element; LTR, long terminal repeat; SINE, short interspersed nuclear element.
a, Repeat annotations across each ape sex chromosome are depicted as a percentage of total nucleotides. Previously uncharacterized human repeats derived from the CHM13 genome analyses are shown in teal. Newly defined satellites (Methods) are depicted in light orange. b, The amount of DNA on each sex chromosome comprising canonical satellites, with each satellite represented by a different colour. LINE, long interspersed nuclear element; LTR, long terminal repeat; SINE, short interspersed nuclear element.
Fig. 5: Centromeres on ape sex chromosomes. a, Left, active alpha satellite suprafamilies (SFs) on the primate phylogenetic tree. Active centromeres in each chromosome have different higher-order repeats in chromosome-specific organization and similar repeats in pan-chromosomal organization. Right, centromeres for each branch (not to scale) with alpha satellite suprachromosomal family composition of the active core indicated in the middle and of the dead flanking layers on the sides. Each branch has one or more alpha satellite suprachromosomal family fewer than in African apes but may also have layers not shared with human (indicated by hues of the same colour). The African ape centromere cores are shown as horizontal bars of SF1–SF3 as each chromosome usually has one alpha satellite suprafamily, which differs with each chromosome. b, The UCSC Genome Browser tracks of alpha satellite suprafamily composition of centromere cores and flanks for cenY and cenX (not to scale). CenX is surrounded by stable vestigial layers (that is, the remnants of ancestral centromeres), whereas cenY has a ‘naked’ centromere devoid of such layers. Thin grey lines under the tracks show overlaps with segmental duplications. In gorilla cenX, SF3 was replaced by SF2 and then by SF1 (see details in Supplementary Note 7).
a, Left, active alpha satellite suprafamilies (SFs) on the primate phylogenetic tree. Active centromeres in each chromosome have different higher-order repeats in chromosome-specific organization and similar repeats in pan-chromosomal organization. Right, centromeres for each branch (not to scale) with alpha satellite suprachromosomal family composition of the active core indicated in the middle and of the dead flanking layers on the sides. Each branch has one or more alpha satellite suprachromosomal family fewer than in African apes but may also have layers not shared with human (indicated by hues of the same colour). The African ape centromere cores are shown as horizontal bars of SF1–SF3 as each chromosome usually has one alpha satellite suprafamily, which differs with each chromosome. b, The UCSC Genome Browser tracks of alpha satellite suprafamily composition of centromere cores and flanks for cenY and cenX (not to scale). CenX is surrounded by stable vestigial layers (that is, the remnants of ancestral centromeres), whereas cenY has a ‘naked’ centromere devoid of such layers. Thin grey lines under the tracks show overlaps with segmental duplications. In gorilla cenX, SF3 was replaced by SF2 and then by SF1 (see details in Supplementary Note 7).
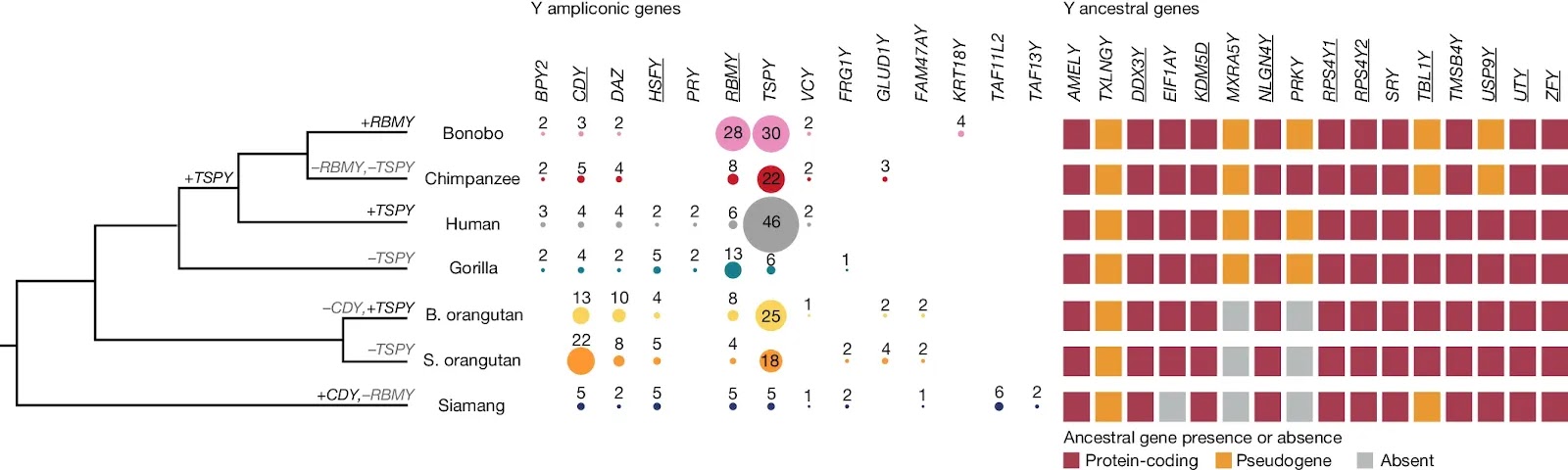
Fig. 6: Gene evolution on the Y chromosome. Significant gains and losses in ampliconic gene copy number (Supplementary Note 10) are shown on the phylogenetic tree. Copy numbers of ampliconic genes are indicated with numbers and by circle size; no circle indicates absence of annotated protein-coding copies. Presence, pseudogenization or absence (that is, deletion) of ancestral (X-degenerate) genes are shown by squares of different colours. Genes showing signatures of purifying selection (Methods) are underlined. XKRY was found to be a pseudogene in all species studied and is therefore not shown. The protein-coding status of PRY was confirmed for human8, and we found evidence of expression of a similar transcript in gorilla (Supplementary Table 36b). The RBMY gene family harboured two distinct gene variants, each present in multiple copies in Pongo (Supplementary Fig. 19).
Significant gains and losses in ampliconic gene copy number (Supplementary Note 10) are shown on the phylogenetic tree. Copy numbers of ampliconic genes are indicated with numbers and by circle size; no circle indicates absence of annotated protein-coding copies. Presence, pseudogenization or absence (that is, deletion) of ancestral (X-degenerate) genes are shown by squares of different colours. Genes showing signatures of purifying selection (Methods) are underlined. XKRY was found to be a pseudogene in all species studied and is therefore not shown. The protein-coding status of PRY was confirmed for human8, and we found evidence of expression of a similar transcript in gorilla (Supplementary Table 36b). The RBMY gene family harboured two distinct gene variants, each present in multiple copies in Pongo (Supplementary Fig. 19).
Makova, K.D., Pickett, B.D., Harris, R.S. et al.
The complete sequence and comparative analysis of ape sex chromosomes.
Nature (2024). https://doi.org/10.1038/s41586-024-07473-2
Copyright: © 2024 The authors.
Published by Springer Nature Ltd. Open access.
Reprinted under a Creative Commons Attribution 4.0 International license (CC BY 4.0)




- The mammalian sex chromosomes began to diverge from a pair of autosomes about 170 million years before the mythical 'Creation Week'.
- The great Ape sex chromosomes began to diverge 17 million years before the mythical 'Creation Week'.
- Chimpanzees, bonobos and humans have almost the same size X chromosomes (154 million nucleotides, compared to the Gorilla's 178 million)
- There is almost as much difference between the X chromosomes of the two species of Orangutan(98.6% alignment) than between humans and chimpanzees (98%) and only a little more than between humans and bonobos (97.7%).
- The Y Chromosomes have evolved by loss of genetic information, mutations and other mechanisms that creationist dogma declares to be invariably fatal, but the conserved sequences of Y chromosome DNA show a high degree of consistency, showing their common origin from an ancestral chromosome.
- The dependence of the researchers on the Theory of Evolution to explain and predict the results of these comparisons with no hint that a supernatural explanation is needed because the TOE is no longer fit for purpose.
What Makes You So Special? From The Big Bang To You

Ten Reasons To Lose Faith: And Why You Are Better Off Without It


No comments:
Post a Comment
Obscene, threatening or obnoxious messages, preaching, abuse and spam will be removed, as will anything by known Internet trolls and stalkers, by known sock-puppet accounts and anything not connected with the post,
A claim made without evidence can be dismissed without evidence. Remember: your opinion is not an established fact unless corroborated.