
Pleurophyllum speciosum - Campbell Island.
Credit Phil Garnock-Jones.
Creationists try to get round the absurdity of the Bible myth which has two (or seven) of every species being packed into a wooden boat small enough to survive turbulent seas for a year, by introducing a new element to the myth that their god forgot to include - that there were just two (or seven) of each 'kind' and all of them underwent a period of warp-speed evolution (that non-one seemed to have noted) with several new species popping into existence each generation to give the many millions of known terrestrial species we have today.
Understandably, creationists are reticent to put any numbers on their claim. They won't say how many different 'kinds' there were on the boat, how many new species arose at each generation and for how long this period of fantastical speciation lasted. Nor will they define 'kind' in any meaningful way that matches any recognisable taxon. I have even been told it can mean 'animal kind' and 'plant kind'. It seems to vary according to the needs of the argument.
And they won't say why some 'kinds' have just one or two species while others have hundreds, or in the case of the Asteraceae family of plants, some 34,000 distinct species, so some must have been speciating much faster then other while some hardly bothered if at all.
In the later case, we now have a substantial database compiled by a team at the Naturalis Biodiversity Center, Leiden, The Netherlands which catalogues all 34,000 different species with their geographical distribution, showing how they radiated and diversified into new species, colonising isolated islands and, like Darwin's finches, radiating into different species on each island in an archipelago.
Tell me all about the Asteraceae family of plants and what is known of their adaptive radiation and colonisation of the planet, including the major taxons within the family. The Asteraceae (or Compositae) family is one of the largest and most diverse families of flowering plants, with over 32,000 species across more than 1,900 genera. They are commonly known as the aster, daisy, composite, or sunflower family. Asteraceae are widespread across most terrestrial ecosystems, colonizing various habitats, and have played a significant role in the flora of many regions worldwide due to their remarkable adaptive radiation.The Naturalis team have just published their findings, open access, in Nature Communications and announced it in a press release:
Key Characteristics
Asteraceae plants are distinguished by their composite flower heads, which consist of many smaller flowers (florets) grouped together. These flower heads (inflorescences) mimic a single large flower, enhancing pollinator attraction. The family has significant diversity in life forms, including herbs, shrubs, and small trees.
- Flower structure: The inflorescence is a capitulum, where multiple florets are arranged on a disc-like receptacle. The florets can be either disk florets (tubular) or ray florets (petal-like structures).
- Fruit: The fruit is typically an achene, a dry, one-seeded fruit that does not open to release the seed.
- Pappus: Many members of the family have a pappus attached to the seeds, facilitating wind dispersal.
Major Taxa within Asteraceae
The family is divided into several subfamilies and tribes, with the most prominent being:
- Asteroideae: The largest subfamily, comprising more than 70% of the species, it includes many well-known genera like Helianthus (sunflower), Aster, Solidago (goldenrods), and Bellis (daisies).
- Cichorioideae: This subfamily includes many plants with milky sap and often showy, strap-shaped ray florets. Some well-known members include Taraxacum (dandelions), Lactuca (lettuce), and Cichorium (chicory).
- Carduoideae: This subfamily includes thistles and their relatives, such as Cirsium and Carduus.
- Mutisioideae: One of the earliest diverging lineages of the family, with members often adapted to more tropical or subtropical climates. Examples include Mutisia and Chaetanthera.
- Barnadesioideae: A small, basal subfamily endemic to South America, this group represents some of the most primitive members of Asteraceae, such as Barnadesia.
Adaptive Radiation and Colonization
Asteraceae are a prime example of adaptive radiation, especially in how they have colonized a wide range of ecological niches. This radiation is attributed to several evolutionary advantages:
- Morphological versatility: Asteraceae species have evolved different growth forms (from herbs to shrubs and trees), enabling them to occupy diverse habitats, from alpine to desert and tropical environments.
- Specialized inflorescence: The composite nature of their flower heads allows for more efficient pollination, as each inflorescence mimics a larger flower, attracting a broader range of pollinators like bees, butterflies, and flies.
- Efficient seed dispersal mechanisms: Many species possess wind-dispersed seeds equipped with a pappus, which increases their potential to spread over vast areas.
- Polyploidy: A high incidence of polyploidy (multiple sets of chromosomes) in Asteraceae has contributed to their speciation and adaptive potential, enabling them to evolve rapidly in response to new environmental challenges.
Global Distribution
Asteraceae are found on every continent except Antarctica, and their success is attributed to their ability to adapt to a wide range of climates. They are particularly dominant in temperate and arid regions, but also have significant representation in tropical environments. Their ability to colonize disturbed areas has also allowed them to thrive in many human-modified landscapes.
Ecological and Economic Importance
Asteraceae species play vital roles in ecosystems as primary food sources for pollinators and herbivores. Economically, many Asteraceae plants are cultivated as food crops (Lactuca, Cynara), ornamentals (Helianthus, Tagetes), and medicinal plants (Echinacea, Artemisia).
In summary, the Asteraceae family is an extraordinary example of adaptive radiation and global colonization. Their evolutionary success is driven by their inflorescence structure, seed dispersal adaptations, and ecological flexibility, allowing them to occupy diverse habitats across the planet.
Daisies in isolation:
the astonishing diversity of botanical family Asteraceae on oceanic islands
A new study reveals that Asteraceae, the botanical family that includes daisies and sunflowers, forms the most diverse group of plants on dozens of islands worldwide. The humble daisies may turn out to be a gold mine for understanding why and how new species evolve in the world’s most isolated environments - that is, if we manage to study them before they go extinct.
The most diverse plant family Tenerife Daisy, Argyranthemum tenerifaeCredit: Lizzie Roeble.
Tenerife Daisy, Argyranthemum tenerifaeCredit: Lizzie Roeble.
The research team, among which Naturalis researchers Luis Valente and Lizzie Roeble, has created a comprehensive global database of island Asteraceae species, revealing that it is the most diverse plant family on oceanic islands. The family consists of around 34,000 species with a wide global distribution. They include familiar names such as artichoke, chamomile, dahlia, and lettuce, daisies, and sunflowers. It was a surprise that more isolated islands tend to host higher numbers of unique species. With over 6,000 species analyzed, nearly 60% are found only on islands.
The treasure chest of tropic islands
Asteraceae have undergone rapid evolutionary changes on islands, resulting in species that often differ significantly from their mainland relatives. This evolutionary process is known as species radiation. Radiations often lead to striking differences in size, shape, and habitat preferences among the new species.
From the modest stickseeds (Bidens) of the Pacific Islands, to the alien-looking and highly endangered silverswords of Hawai’i, to the giant daisy trees (Scalesia) of the Galápagos. Although species radiations are more famously documented in animals, such as Darwin’s finches in the Galápagos, this research indicates that Asteraceae may host a record number of radiations on islands worldwide. Many of these radiations are yet to be formally discovered, mostly located on tropical islands such as the Caribbean, Madagascar, and New Guinea.
A surprising count of radiations
Botanists had long suspected that the daisy family had evolved in remarkable ways on islands, but our study reveals that the extent of evolutionary innovation in this family may be much greater than previously imagined.
Lizzie Roeble, lead author
Naturalis Biodiversity Center
Darwinweg, Leiden, The Netherlands.
Global hotspot for evolution
This discovery opens up new possibilities for understanding the mechanisms driving plant diversification. Many of these species, however, are critically endangered, with only a few individuals surviving in remote and often inaccessible island habitats. Given their restricted distributions and unique evolutionary history, the protection of these species is crucial. Island ecosystems harbor not only incredible diversity but also highly specialized species.
Island conservation is urgent
The study reminds us of the vulnerable biodiversity on islands. Many of these species are found in highly localized, inaccessible areas and face threats from habitat loss, climate change, and human activities. The daisy family has an incredible evolutionary potential. Their conservation is essential in preventing these unique species from disappearing forever.
More informationRead the original article ‘Island biogeography of the megadiverse plant family Asteraceae’ by Roeble, L., van Benthem, K.J., Weigelt, P., Kreft, H., Knope, M.L., Mandel, J.R., Vargas, P., Etienne, R.S., Valente, L (2024)

AbstractTucked away in the body of the paper is the revelation that an initial colonisation of the Hawaiian islands (from the North American mainland) was some 5 million years ago, since when the colonists has radiated into 33 species in 3 genera. Another colonisation some 1.8 million years ago resulted in a single genus of 20 species. similar colonisation by a single species followed by radiating speciation have occurred on the Galapagos Islands, the Canary Islands, the Juan Fernández Islands (30 species in 4 genera).
The megadiverse plant family Asteraceae forms an iconic component of island floras including many spectacular radiations, but a global picture of its insular diversity is lacking. Here, we uncover the global biogeographical and evolutionary patterns of Asteraceae on islands to reveal the magnitude and potential causes of their evolutionary success. We compile a global checklist of Asteraceae species native and endemic to islands and combine it with macroecological analyses and a phylogenetic review of island radiations. Asteraceae have a global distribution on islands, comprising approximately 6,000 native island species, with 58% endemics. While diversity of the family on islands is lower than expected given its overall diversity, Asteraceae are the most diverse family on oceanic islands, suggesting an exceptional ability to thrive in isolation. In agreement with island biogeography predictions, native Asteraceae diversity increases with area and decreases with isolation, while endemism increases with both. We identify 39 confirmed island radiations and 69 putative radiations, exceeding numbers for other iconic insular groups. Our results reveal Asteraceae offer immense potential for research in ecology and evolution, given their close tracking of island biogeography expectations, large number of both species and radiations, cosmopolitan distribution, and numerous undiscovered radiations.
Introduction
The top ten most diverse plant families make up 43% of angiosperm species1. Understanding the distribution and drivers of diversity of these large families is thus a crucial step towards explaining the success of flowering plants in general. Key biogeographical settings for exploring the patterns and processes that shape angiosperm diversity are islands. Due to their distinct boundaries, global distribution, and replication, island systems have played a crucial role in the development of key evolutionary and ecological theories2,3,4,5,6,7,8. The geographic isolation and unique habitats found on islands have given rise to remarkable angiosperm biodiversity that is often characterized by high levels of endemism9,10, adaptive radiations11,12, paleoendemism13,14, and repeated evolution of convergent traits15,16.
While islands are valuable natural laboratories for studying plant diversity, global-scale data on the distribution of major plant families on islands are only starting to emerge. Recent global studies have explored biodiversity patterns for a few major families and lineages on islands17,18, factors impacting the assembly of island floras19,20,21,22, and traits associated with insular diversity23,24,25,26. These studies reveal how links between island features (e.g., area, isolation, age, and climate), functional traits (e.g., insular woodiness), and biogeographical rates (e.g., colonization, speciation, and extinction) are important determinants of the number of native and endemic species of flowering plants on islands, whilst suggesting that evolutionary success on islands may not necessarily mirror that found on continents27.
Out of all plant families, arguably the one most often associated with evolutionary success on islands is the most diverse family of all—Asteraceae. Commonly known as the daisy or sunflower family, Asteraceae (Compositae) boasts the greatest species number of any plant family in the world, with an estimated 32,000–34,000 species1,28,29. Species of this family are native to every continent except Antarctica and are found in a wide range of habitats, but are most abundant in dry and semi-arid habitats and in Mediterranean-type ecosystems, deserts, grasslands, and mountains30. Members of the family show great variation in growth habits and range from small annual herbs to woody perennial shrubs, lianas, and trees. Additionally, some species exhibit specialized habits such as cushion forms, succulents, and even, in rare cases, are epiphytic and aquatic plants.
On islands, Asteraceae are thought to be remarkably diverse, and often form an iconic component of insular floras of both continental and oceanic origin. For instance, it is the most species-rich family on the remote Juan Fernández Archipelago31 with 30 native species and four genera endemic to the islands, and is among the top five most diverse families on the large continental island of Madagascar32. Additionally, the family has high levels of endemism on oceanic islands: a study by Lenzner et al.33 compiled diversity data on major plant families across 14 oceanic archipelagos and found that Asteraceae had the highest number of single-island endemics for the oceanic islands considered in the study. Their success in dispersal, establishment, and diversification on islands has been suggested to result from a combination of intrinsic factors3,34,35: Asteraceae possess unique fruit morphology that aids in long-distance dispersal36,37; their head-like inflorescence (capitulum) often attracts generalist pollinators; and they are capable of several breeding systems that may favor establishment on islands. Many island species are self-compatible23, while species in several island lineages are functionally self-incompatible with the capacity to self-seed and a genetic system (i.e., sporophytic self-incompatibility) that aids both in the establishment of small populations and the retention of genetic diversity after arrival38.
In addition to a high native and endemic species richness on islands, Asteraceae are known for their presumed propensity to radiate (that is, to undergo cladogenesis in situ on islands at relatively fast rates). Two recent studies, one reviewing adaptive radiations across flowering plants39 and another focused on adaptive radiations on oceanic islands across all taxonomic groups40, both found Asteraceae to be overrepresented in terms of adaptive radiations compared to other clades. Indeed, the family provides numerous examples of spectacular island radiations: Scalesia on the Galápagos Islands41, the woody Sonchus alliance on the Canary Islands42, Dendroseris on the Juan Fernández Islands43,44. One of the textbook examples of adaptive radiation on islands is the Asteraceae silversword alliance of Hawai‘i, a clade of 33 species in three endemic genera (Argyroxiphium, Dubautia, Wilkesia), which evolved from a common ancestor that colonized Hawai‘i by a long-distance dispersal event from North America around 5 million years ago (Mya), and which exhibit high diversity in morphology and ecological adaptation45,46,47. Another notable example is the Hawaiian Bidens. The monophyletic 20 species of Bidens endemic to Hawai‘i originated from a single colonization event c. 1.8 Mya, having thereafter radiated across the archipelago, occupying a wide variety of different habitats including sand dunes, lava fields, rainforests, and wetland bogs, and have the highest rates of speciation per unit area documented for any island plant radiation to date48,49.
An increasing number of phylogenetic studies focusing on selected island clades of Asteraceae from specific islands or archipelagos41,47,50,51,52 are providing insight into the potential drivers of diversification in those Asteraceae groups. One hypothesis is that the high diversity of Asteraceae on islands results from a combination of high continental diversity, high rates of long-distance dispersal, and overall high rates of in situ speciation that well exceed extinction rates (consistent with the high net diversification rates observed in continental Asteraceae)53,54,55,56.
While it is assumed from the above examples that Asteraceae are highly diverse on islands and have a propensity to radiate, in fact, a complete global picture of the diversity and distribution of the family is yet to be assembled. Furthermore, an assessment of Asteraceae’s potential to radiate across islands globally is still lacking, because previous studies focused solely on confirmed adaptive radiations and/or on oceanic islands, and thus the magnitude of island radiations within the family is unknown.
In this work, we compile a global checklist of island Asteraceae and use this to answer four key questions: (1) How does the island species richness of Asteraceae compare with that of other flowering plant families? (2) How is island Asteraceae diversity distributed across space and major clades of the family? (3) What are the environmental and biogeographical drivers of native and endemic insular diversity on islands? (4) How many island radiations have occurred within Asteraceae and are there commonalities between radiations? We show that Asteraceae have a truly global distribution on islands, with 6135 native island species, 58% of which are endemic. Our findings reveal that while the diversity of the family on islands is lower than expected given their overall diversity, Asteraceae are the most diverse family on oceanic islands, suggesting an exceptional ability to thrive in isolation. Moreover, diversity patterns in Asteraceae follow classic island biogeography theory, with area and isolation being the strongest predictors of species richness and endemism, and the numerous confirmed and understudied radiations highlight the family’s potential as an ideal study system in ecology and evolution.Fig. 1: The ten most diverse angiosperm families on islands. Species diversity of these families on A all islands and B oceanic islands compared to the null expectation. The left panels rank the ten most diverse angiosperm families on islands for native (blue and purple points) and endemic (green and orange points) species. Families are ranked in descending order by the number of native species. The right panel compares the observed number of native island species per family (points) to the null expectation of island diversity (histogram). Families for which the observed number of species is lower than the null expectation are highlighted with a red point, and those above the null expectation in black. Source data are provided in the Source Data file. The global diversity of each family is listed in Supplementary Table 2.
Species diversity of these families on A all islands and B oceanic islands compared to the null expectation. The left panels rank the ten most diverse angiosperm families on islands for native (blue and purple points) and endemic (green and orange points) species. Families are ranked in descending order by the number of native species. The right panel compares the observed number of native island species per family (points) to the null expectation of island diversity (histogram). Families for which the observed number of species is lower than the null expectation are highlighted with a red point, and those above the null expectation in black. Source data are provided in the Source Data file. The global diversity of each family is listed in Supplementary Table 2.
Fig. 2: Geographical diversity patterns of island Asteraceae. A Global distribution of Asteraceae across all island types. The shape of the marker represents the island type (i.e., continental or oceanic), the size represents the number of species, and the color indicates the number of endemics, with gray meaning no island endemics. B Global distribution of Asteraceae on archipelagos. Source data are provided in Supplementary Data 3.
A Global distribution of Asteraceae across all island types. The shape of the marker represents the island type (i.e., continental or oceanic), the size represents the number of species, and the color indicates the number of endemics, with gray meaning no island endemics. B Global distribution of Asteraceae on archipelagos. Source data are provided in Supplementary Data 3.
Fig. 3: Island diversity within the subfamilies and tribes of Asteraceae. A Time-calibrated molecular phylogeny of the tribes and major clades within Asteraceae from ref. 54. Tribes are colored by subfamily classification. B The number of species native to islands (dark bar) compared to the overall number of species globally (light bar) in each tribe. The percentage of native island species to total species globally is specified next to each tribe. Illustrations highlight clades with high island diversity: (1) Argyroxiphium sandwicense endemic to Hawai‘i, Madieae; (2) Commidendrum rugosum endemic to Saint Helena, Astereae; (3) Abrotanella inconspicua endemic to New Zealand, Senecioneae; (4) Distephanus populifolius endemic to Mauritius, Distephaneae; (5) Anastraphia ilicifolia endemic to Cuba, Gochnatiaeae. Plant illustrations by Lizzie Roeble, and originally featured in the CAPITULUM. The Asteraceae phylogeny is adapted from Mandel, J. R. et al. A fully resolved backbone phylogeny reveals numerous dispersals and explosive diversifications throughout the history of Asteraceae. Proc. Natl. Acad. Sci. USA 116, 14083–14088 (2019). The phylogeny and plant illustrations are released under a Creative Commons Attribution-NonCommercial-NoDerivs 4.0 International license (https://creativecommons.org/licenses/by-nc-nd/4.0/). Source data are provided in the Source Data file.
A Time-calibrated molecular phylogeny of the tribes and major clades within Asteraceae from ref. 54. Tribes are colored by subfamily classification. B The number of species native to islands (dark bar) compared to the overall number of species globally (light bar) in each tribe. The percentage of native island species to total species globally is specified next to each tribe. Illustrations highlight clades with high island diversity: (1) Argyroxiphium sandwicense endemic to Hawai‘i, Madieae; (2) Commidendrum rugosum endemic to Saint Helena, Astereae; (3) Abrotanella inconspicua endemic to New Zealand, Senecioneae; (4) Distephanus populifolius endemic to Mauritius, Distephaneae; (5) Anastraphia ilicifolia endemic to Cuba, Gochnatiaeae. Plant illustrations by Lizzie Roeble, and originally featured in the CAPITULUM. The Asteraceae phylogeny is adapted from Mandel, J. R. et al. A fully resolved backbone phylogeny reveals numerous dispersals and explosive diversifications throughout the history of Asteraceae. Proc. Natl. Acad. Sci. USA 116, 14083–14088 (2019). The phylogeny and plant illustrations are released under a Creative Commons Attribution-NonCommercial-NoDerivs 4.0 International license (https://creativecommons.org/licenses/by-nc-nd/4.0/). Source data are provided in the Source Data file.
Fig. 5: Summary of island radiations within Asteraceae.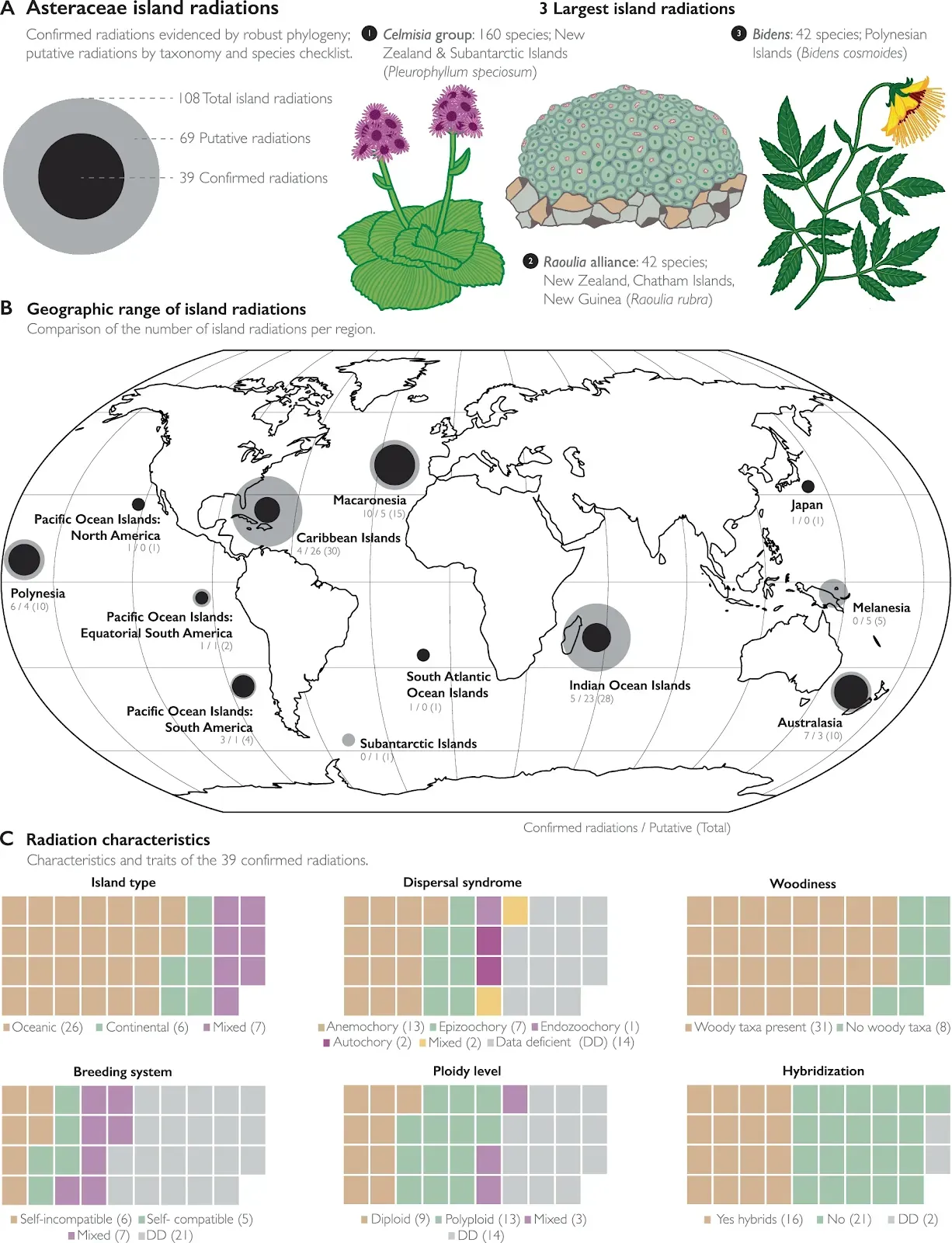 A Overview of the number of total, confirmed, and putative insular radiations within Asteraceae. The confirmed radiations have been evidenced by robust phylogenetic work and are represented by the black circles, and the putative radiations have been identified based on taxonomy and the island Asteraceae checklist and are represented by the gray circles (see methods for details on assessment criteria). Illustrations of species within the three largest island radiations. B Map compares the number of radiations between regions. In cases where a radiation is distributed across multiple regions, it is included in the region where the most species diversity is located. Several island regions had no radiations (Mediterranean Islands, Micronesia). C The waffle charts summarize characteristics and traits of the confirmed radiations, where a single radiation is represented by one square. Traits were scored at the radiation level, and if there are multi-states in the radiation it is captured with the ''Mixed” category. Plant illustrations by Lizzie Roeble, originally featured in the CAPITULUM and released under a Creative Commons Attribution-NonCommercial-NoDerivs 4.0 International license (https://creativecommons.org/licenses/by-nc-nd/4.0/). Full review of island radiations in Supplementary Data 4.
A Overview of the number of total, confirmed, and putative insular radiations within Asteraceae. The confirmed radiations have been evidenced by robust phylogenetic work and are represented by the black circles, and the putative radiations have been identified based on taxonomy and the island Asteraceae checklist and are represented by the gray circles (see methods for details on assessment criteria). Illustrations of species within the three largest island radiations. B Map compares the number of radiations between regions. In cases where a radiation is distributed across multiple regions, it is included in the region where the most species diversity is located. Several island regions had no radiations (Mediterranean Islands, Micronesia). C The waffle charts summarize characteristics and traits of the confirmed radiations, where a single radiation is represented by one square. Traits were scored at the radiation level, and if there are multi-states in the radiation it is captured with the ''Mixed” category. Plant illustrations by Lizzie Roeble, originally featured in the CAPITULUM and released under a Creative Commons Attribution-NonCommercial-NoDerivs 4.0 International license (https://creativecommons.org/licenses/by-nc-nd/4.0/). Full review of island radiations in Supplementary Data 4.
Roeble, L., van Benthem, K.J., Weigelt, P. et al.
Island biogeography of the megadiverse plant family Asteraceae.
Nat Commun 15, 7276 (2024). https://doi.org/10.1038/s41467-024-51556-7
Copyright: © 2024 The authors.
Published by Springer Nature Ltd. Open access.
Reprinted under a Creative Commons Attribution 4.0 International license (CC BY 4.0)




To explore this a little further, here is what ChatGPT4o has provided:
Do you have information on which oceanic archipelagoes have been colonised with approximate dates and the number of descendant genera and species? Yes, Asteraceae have successfully colonized many oceanic archipelagos, showing significant adaptive radiation in these isolated environments. Though dates of colonization are often difficult to determine precisely, estimates are made using molecular phylogenetics, fossil records, and geological dating of islands. Below are some notable examples of oceanic archipelagos colonized by Asteraceae, along with estimated dates and the number of descendant genera and species where known.Rapid and wide-ranging though these patterns of colonisation and evolutionary radiations into multiple genera and species are, by biological evolutionary standards, occurring mostly in 1-10 million years, this is slower by several orders of magnitude than creationists like to imagine was possible when the animals left the Ark in their genocidal flood myth.
- Hawaiian Archipelago
The Hawaiian Islands are among the most studied cases of Asteraceae radiation. The silversword alliance is a classic example of adaptive radiation, where a single ancestral colonizer evolved into a diverse group of species occupying a wide range of ecological niches, from coastal shrublands to alpine zones.
- Colonization Date: Estimated around 5 million years ago.
- Key Genera: Dubautia, Argyroxiphium, Wilkesia (collectively known as the silversword alliance).
- Number of Descendant Species: Around 30 species within the silversword alliance.
- Galápagos Archipelago
The genus Scalesia in the Galápagos is often referred to as the “Darwin’s finches of the plant world” due to its rapid adaptive radiation. Species in this genus have evolved to thrive in a variety of habitats, including dry lowlands and humid highlands.
- Colonization Date: Estimated 1–4 million years ago.
- Key Genera: Scalesia.
- Number of Descendant Species: Around 15 species.
- Juan Fernández Islands (Chile)
The genus Dendroseris, endemic to the Juan Fernández Islands, has evolved from an ancestral Asteraceae colonizer into several woody species. These plants display characteristics well adapted to the island's unique environments, including cloud forests and rocky cliffs.
- Colonization Date: Estimated around 2–3 million years ago.
- Key Genera: Dendroseris.
- Number of Descendant Species: About 11 species.
- Canary Islands
The Canary Islands, with their range of climatic zones, have seen considerable diversification in the Asteraceae family. Sonchus species have radiated significantly in this archipelago, evolving into a wide array of forms, from herbaceous plants to small trees.
- Colonization Date: Estimated around 5–10 million years ago.
- Key Genera: Sonchus, Launaea, Tolpis, Sventenia.
- Number of Descendant Species: Around 30–50 species.
- New Zealand
New Zealand has a rich diversity of Asteraceae, with genera such as Celmisia and Olearia contributing significantly to its alpine and coastal ecosystems. Adaptive radiation in these genera has led to a wide variety of forms, including shrubs, trees, and cushion plants.
- Colonization Date: Estimated 1–2 million years ago.
- Key Genera: Celmisia, Olearia, Brachyglottis.
- Number of Descendant Species: Over 200 species across multiple genera.
- Madagascar
Madagascar has experienced significant radiation of Asteraceae, particularly in the genus Helichrysum. These species occupy a range of ecological zones, from dry forests to high-altitude heathlands.
- Colonization Date: Estimated 10–15 million years ago.
- Key Genera: Helichrysum, Vernonia.
- Number of Descendant Species: Over 150 species.
- Society Islands (French Polynesia)
In the Society Islands, the genus Bidens has radiated into various forms, including woody shrubs, adapting to a variety of island habitats. This genus is well-known for its presence in both tropical and temperate island systems.
- Colonization Date: Estimated around 1–2 million years ago.
- Key Genera: Bidens.
- Number of Descendant Species: Around 10–15 species.
- Mascarenes (Réunion, Mauritius, Rodrigues)
The genus Psiadia is notable in the Mascarenes, where it has diversified into numerous species that have adapted to different ecological niches on these volcanic islands.
- Colonization Date: Estimated 3–5 million years ago.
- Key Genera: Psiadia.
- Number of Descendant Species: Around 25 species.
Patterns of Colonization and Radiation
- Single or Multiple Colonization Events: In many archipelagos, Asteraceae colonization is thought to have resulted from a single ancestral species, which then underwent rapid adaptive radiation. In some cases, like the Hawaiian Islands, multiple colonization events may have occurred.
- Ecological Diversification: Asteraceae species on islands have diversified to fill a variety of niches, including coastal, montane, and arid habitats. Many have evolved unique traits, such as woodiness in typically herbaceous genera (e.g., Sonchus in the Canary Islands).
- Reproductive Isolation and Speciation: Due to geographic isolation, the genetic divergence of populations has led to high species endemism on oceanic islands. Island populations often evolve specialized pollination strategies, seed dispersal mechanisms, and life forms suited to their environments.
Conclusion
Oceanic archipelagos have provided Asteraceae with opportunities for incredible adaptive radiation. These islands, isolated from mainland floras, allow for rapid speciation, leading to the evolution of diverse forms from a few ancestral species. Molecular dating, combined with island geology and the fossil record, has allowed scientists to estimate the timing of these colonization events, though exact dates are still subject to ongoing research.
And of course, like almost all the history of life on Earth, this all happened in the vast pre-'Creation Week' period before their god reputedly made a small flat planet with a dome over it, out of nothing, somewhere in the Middle East.
Of course, creationists will settle for the stock answer, "God did it!" and feel satisfied with not knowing how it really happened and shoehorning their little god into another shrinking gap. Scientists, on the other hand, will try to work out and understand what it is about the Asteraceae that makes it possible for them to disperse so widely, then speciate rapidly to fit into available ecological niches, and so evict their little god from the cosy god-shaped gap that creationists fashioned for it out of their ignorance.
What Makes You So Special? From The Big Bang To You

How did you come to be here, now? This books takes you from the Big Bang to the evolution of modern humans and the history of human cultures, showing that science is an adventure of discovery and a source of limitless wonder, giving us richer and more rewarding appreciation of the phenomenal privilege of merely being alive and able to begin to understand it all.
Available in Hardcover, Paperback or ebook for Kindle
Ten Reasons To Lose Faith: And Why You Are Better Off Without It

This book explains why faith is a fallacy and serves no useful purpose other than providing an excuse for pretending to know things that are unknown. It also explains how losing faith liberates former sufferers from fear, delusion and the control of others, freeing them to see the world in a different light, to recognise the injustices that religions cause and to accept people for who they are, not which group they happened to be born in. A society based on atheist, Humanist principles would be a less divided, more inclusive, more peaceful society and one more appreciative of the one opportunity that life gives us to enjoy and wonder at the world we live in.
Available in Hardcover, Paperback or ebook for Kindle

No comments:
Post a Comment
Obscene, threatening or obnoxious messages, preaching, abuse and spam will be removed, as will anything by known Internet trolls and stalkers, by known sock-puppet accounts and anything not connected with the post,
A claim made without evidence can be dismissed without evidence. Remember: your opinion is not an established fact unless corroborated.