

A paper published on 20 May 2026 in Science Advances by a team of palaeontologists led by Scott D. Evans of the American Museum of Natural History, New York, very neatly illustrates the difference between evolutionary biology and creationism. It reports the discovery of a rich new Ediacaran fossil site in the Mackenzie Mountains of Canada’s Northwest Territories, containing fossils that appear earlier in the record, and in deeper-water settings, than current models of the Ediacaran biota had led palaeontologists to expect.
That is not a problem for evolution; it is how science progresses. Unexpected evidence does not destroy a scientific theory merely because it requires a refinement of detail. In this case, the discovery extends the known geographical, ecological and chronological range of part of the Ediacaran biota — the strange, mostly soft-bodied organisms that preceded, and helped set the stage for, the later Cambrian diversification of animal life.
When asked what would falsify the theory of evolution, the evolutionary biologist J.B.S. Haldane is said to have replied, “a fossil rabbit in the Precambrian”. He was making the simple point that evolutionary theory predicts a broad historical sequence: mammals should not appear before vertebrates, vertebrates should not appear before animals, and rabbits should not appear hundreds of millions of years before their ancestors. A genuine rabbit in Precambrian rocks would be an anachronism so extreme that it would call the whole historical framework into question.
But that is not what palaeontologists have found here. The newly reported fossils are not out of sequence; they are exactly the kind of organisms that belong in late Precambrian rocks. The surprise is not that they are in the wrong part of life’s history, but that some of them appear a little earlier, in a wider geographical range, and in somewhat different environments than previously recognised. In other words, the anomaly is chronological and ecological, not evolutionary.
To a creationist, of course, the question of falsification has to be avoided, because the honest answer is deeply uncomfortable. The fossil record as a whole does not show a sudden magical creation of all living things a few thousand years ago. It shows succession: organisms appearing, diversifying, changing and disappearing through vast spans of geological time. The dating of the rocks, using multiple independent geological and radiometric methods, consistently points to an ancient Earth and a long history of life, not to a recent creation week followed by a global flood.
That is why every such discovery is awkward for creationism but routine for science. Fossils are not distributed randomly, as they would be if all life had been created at once and then jumbled together in a recent catastrophe. They occur in a recognisable sequence, constrained by stratigraphy, radiometric dating, comparative anatomy, developmental biology and, for later organisms, genetics. The details are continually revised, but the broad pattern remains overwhelmingly consistent with evolution and wholly inconsistent with Biblical literalism.
By any honest application of the scientific method, that should be enough to falsify the creationist narrative beyond reasonable doubt. That it does not do so for creationists is not because the evidence is weak, but because the conclusion is protected from evidence. When the conclusion is sacred, facts become things to be explained away, misrepresented or ignored.
For evolutionary biologists, however, an unexpected fossil is not an embarrassment to be dismissed, but a clue to be investigated. If the evidence shows that part of the White Sea assemblage was present in Laurentia earlier than previously recognised, and in deeper-water environments, then the scientific response is to refine the model. The theory is not weakened by that process; it is strengthened, because it can absorb new evidence, generate better questions and produce a more accurate account of what happened.
The fossils described in this paper include more than 100 specimens, with several groups not previously recorded from North America, including Dickinsonia, Funisia, Kimberella and Eoandromeda. Some are estimated to be about 567 million years old, overlapping with the older Avalon assemblage and extending the known range of the White Sea assemblage by around 5–10 million years. The researchers also found that these organisms lived in deeper-water settings than had previously been recognised for this assemblage, supporting the idea that some early animal innovations may have begun offshore before spreading into shallower environments.
The Ediacaran Biota and Its Three Broad Assemblages. The Ediacaran Period, roughly 635–541 million years ago, was the final period of the Precambrian, immediately before the Cambrian. It marks one of the most important transitions in the history of life: the change from a world dominated largely by microbes and microbial mats to one in which large, complex, multicellular organisms began to occupy the seafloor.
The organisms of the Ediacaran biota were mostly soft-bodied, which makes their fossilisation unusual and often exceptional. Some were frond-like, some were disc-shaped or quilted, some were tubular, and others appear to have been capable of movement across the seafloor. Many do not fit neatly into modern animal groups, although some may represent early relatives of later animal lineages. Their importance lies not in being “modern” animals, but in showing that complex body plans, ecological interactions and animal-like behaviours were already developing before the Cambrian diversification.
Palaeontologists often divide the Ediacaran fossil record into three broad assemblages: Avalon, White Sea and Nama. These are not separate creations or unrelated faunas, but useful scientific groupings based on differences in age, environment and fossil communities.
- The Avalon assemblage, known especially from Newfoundland and other sites, is generally the oldest of the three. It is associated with deep-water settings and includes many frond-like organisms that appear to have lived attached to the seafloor. These communities show that large, complex life was already established in marine environments long before the Cambrian.
- The White Sea assemblage, named after fossil localities near the White Sea in Russia, is younger and typically more diverse. It includes well-known Ediacaran organisms such as Dickinsonia, Kimberella and Funisia. Some of these fossils have been interpreted as evidence of movement, feeding behaviour and possibly early forms of sexual reproduction, making this assemblage especially important for understanding the emergence of recognisably animal-like lifestyles.
- The Nama assemblage, known from sites such as Namibia, is generally the youngest of the three and approaches the boundary with the Cambrian. It includes organisms with more resistant or skeletal elements, hinting at the ecological and evolutionary changes that would become much more obvious during the Cambrian diversification.
The new fossils from the Mackenzie Mountains of north-western Canada are important because they complicate and improve this picture. They show clear evidence of White Sea assemblage organisms in ancient rocks of Laurentia, the ancestral core of North America, and some of them appear around 5–10 million years earlier than previously documented examples. They were also found in deposits interpreted as deeper-water settings than expected for this assemblage.
That does not overturn evolution; it refines it. The discovery suggests that some Ediacaran organisms were more widespread, appeared earlier and occupied a broader range of environments than palaeontologists had previously recognised. It is therefore a good example of how science works: not by protecting a fixed story from inconvenient facts, but by adjusting the story when new evidence demands it.
The paper’s publication was accompanied by a news item from the American Museum of Natural History:
Fossil Trove Expands Range of Squishy Early Animals
Hundreds of millions of years before the age of dinosaurs, during a time known as the Ediacaran, life existed mostly in the oceans, and the seafloor was covered with soft organisms unlike anything alive today.
With shapes ranging from delicate fern-like fronds to segmented blobs, fossils of Ediacaran organisms represent the first direct evidence of multicellular animal life as well as the origins of animal movement and sexual reproduction.
Now, researchers have uncovered a remarkable site in Canada’s Northwest Territories where they have found more than 100 Ediacaran fossils, including six groups never before seen in North America. These fossils are older and found in much deeper water settings than have been previously described. The work is published today in the journal Science Advances.
 Reconstruction of a hypothetical deep-water group from the new fossil site in Canada’s Northwest Territories, based on fossils recovered by the researchers.Alex Boersma.
Reconstruction of a hypothetical deep-water group from the new fossil site in Canada’s Northwest Territories, based on fossils recovered by the researchers.Alex Boersma.For 3 billion years, life on Earth was dominated by microbes. Then, all the sudden, we get these strange-looking marine animals big enough to see and capable of behaviors we would find familiar today. If we want to understand this transition, when life first became large, complex, and unmistakenly animal, this new site has tremendous potential.
Scott D. Evans, lead author
Division of Paleontology (Invertebrates)
American Museum of Natural History
New York, NY, USA.
Ediacaran species are linked to a diverse set of animal groups, including mollusks, nematodes, comb jellies, and cnidarians—a group that spans jellyfish and corals. Because most Ediacaran organisms were soft-bodied, their fossils are rare. Although these fossils have been identified on every continent except Antarctica, our understanding of Ediacaran life primarily comes from just a handful of sites, providing limited insight into this roughly 40-million-year interval in Earth’s history.
Ediacaran life is sorted into three groups, representing different times in the geologic record. These “assemblages” are called Avalon (575-559 million years ago), White Sea (559-550 million years ago), and Nama (550-538 million years ago).
Until this study, fossils from the White Sea assemblage had mostly been found in Russia and Australia. Now, researchers have uncovered clear evidence of this assemblage in North America, in the remote, ancient rocks of Canada’s Mackenzie Mountains.
In addition, the researchers determined that some of these North American fossils are estimated to be about 567 million years old, 5-10 million years older than previously documented White Sea specimens and overlapping with the time of the older Avalon assemblage.
Not only is this new site highly diverse, but also it is from a part of the rock succession where we have previously lacked fossil remains.
Associate Professor Justin Strauss, co-author.
Department of Earth and Planetary Sciences
Dartmouth College
Hanover, NH, USA.
[Jusin Strauss] has been exploring this area for about 15 years. The fossils were found on the traditional lands of the Sahtú Dene and Métis, who provided the research team with guidance and permission to access the site.
Among the finds made for the first time in North America are: In this study, the researchers found clear evidence of the White Sea assemblage in ancient rocks of Canada’s Mackenzie Mountains in the Northwest Territories, on the traditional lands of the Sahtú Dene and Métis, who provided the research team with guidance and permission to access the site.Scott Evans/© AMNH
In this study, the researchers found clear evidence of the White Sea assemblage in ancient rocks of Canada’s Mackenzie Mountains in the Northwest Territories, on the traditional lands of the Sahtú Dene and Métis, who provided the research team with guidance and permission to access the site.Scott Evans/© AMNH
- Dickinsonia, a flat organism resembling a round mat, with a divided circular body that moved around on the sea floor, lacking a mouth, and instead absorbing bacteria and algae through its entire bottom surface;
- an immobile tubular organism called Funisia that lived in clusters and offers the oldest evidence of sexual reproduction in the fossil record, likely with coordinated release of sperm and eggs into the water column like coral;
- Kimberella, an organism with a muscular foot that fed by scraping the sea floor, widely interpreted as an early relative of mollusks and now potentially the oldest fossil bilaterian—the group of animals with distinct front, back, top, and bottom with symmetric left and right sides;
- and Eoandromeda, a possible comb jelly with eight spiral arms
The scientists also discovered that these organisms lived in deeper-water environments than previously recognized for the White Sea assemblage. This finding supports a growing hypothesis that early animals may have originated in offshore, deep-marine settings before expanding into shallower waters over time, the opposite of what is typical of animal evolution after this time. A fossil of Eoandromeda, a possible comb jelly with eight spiral arms.Scott Evans/© AMNH
A fossil of Eoandromeda, a possible comb jelly with eight spiral arms.Scott Evans/© AMNH
We think of the deep ocean as a dark, inhospitable place, but it is also relatively stable, with few fluctuations in things like temperature and oxygen essential to most animal life. This stability may have provided key opportunities to support early animal life.
Scott D. Evans.
Publication:



The paper's significance for understanding the pre-Cambrian evolution of life is also explained in an article in The Conversation by two researchers from Curtin University, Australia: Chris Kirkland, Professor of Geochronology, and Anthony Clarke, Research Fellow in the School of Earth and Planetary Sciences. Their article is reprinted below under a Creative Commons license, reformatted for stylistic consistency.
A new study, published today in Science Advances, adds crucial information to this story. Led by Scott Evans, assistant curator of invertebrate palaeontology at the American Museum of Natural History, it draws on rare 567-million-year-old fossils to show animal evolution may have started far earlier than previously thought.
Some of these soft-bodied and strange animals were shaped like pancakes. Others were more like soft tubes or spirals that pressed into the mud.
We call this time, from about 635 to 538 million years ago, the Ediacaran Period. Do animals from this period represent our ancient ancestors before the Cambrian explosion, which produced most of the basic groups of animals we know today? Or are they failed evolutionary experiments?
To help us answer these questions, we divide the Ediacaran fossil record into three broad chapters: the Avalon, White Sea and Nama assemblages. Each represents a distinctive community of Ediacaran animals that tend to appear in different times and environments.
These chapters help scientists track how early animal life changed from mostly deep-water organisms that were stuck in mud to more diverse shallow-water communities that included animals.
The Avalon assemblage is the oldest chapter, dominated by simple yet strange deeper water organisms. The White Sea assemblage is the middle chapter. It is characterised by larger, more varied animals, including forms such as the famous Dickinsonia, a ribbed, oval organism a bit like a quilted placemat. The Nama assemblage comes last and includes some of the earliest animals with hard shell-like parts.
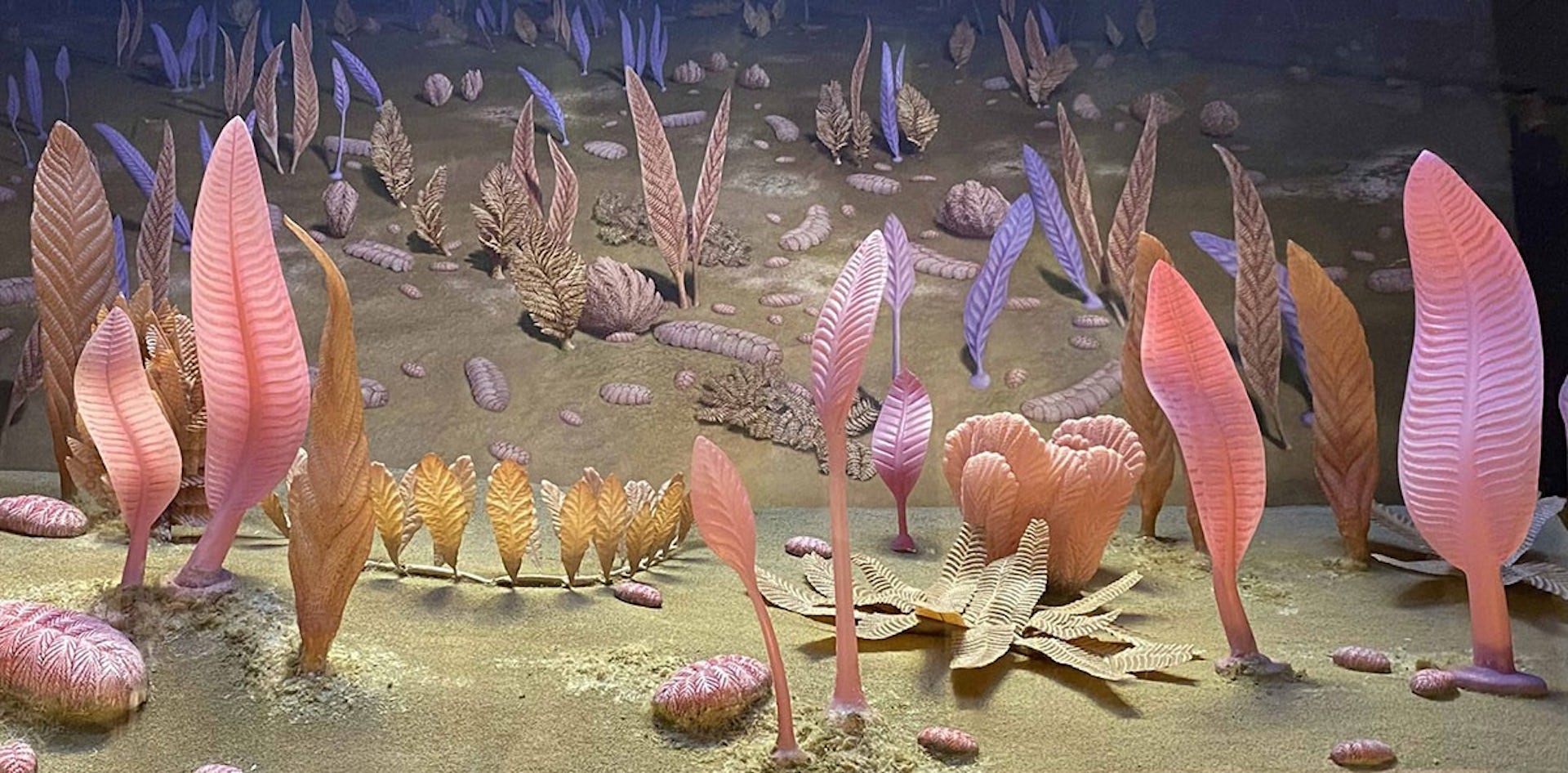
The team behind the new study combined fossil hunting with geological detective work. They collected and photographed fossil-bearing rocks from the remote Mackenzie Mountains in Canada, compared the fossils with other Ediacaran organisms, and studied nearby rocks to reconstruct where and when these animals lived.
Remarkably, several of the fossils, with frond-like forms, segmented and quilted bodies resembled those from the White Sea assemblage. That matters because the White Sea animal community was previously best known from famous sites in Russia and Australia.
The new fossils show that similar communities had also reached the deep waters of Laurentia, the ancient continent that included much of present-day North America.
In early animal evolution, a few million years can matter. The fossil-bearing rocks appear to correlate with nearby layers dated at about 567–566 million years old.
If that correlation is correct, this makes the community considerably older than the classic White Sea assemblage, which is usually placed at about 560–550 million years ago. Their discovery pushes back the timing of some important early animals, including mobile forms such as Dickinsonia.
It also dramatically changes the environmental picture.
White Sea-type fossils are usually associated with shallower marine settings. But these rocks suggest the Canadian animals lived in a deep-water slope environment. Together, that implies these early animal communities were both more geographically widespread and more environmentally flexible than previously recognised.
That raises an intriguing question. Did early animal ecosystems first develop far offshore, in deeper and perhaps more stable marine settings, before later becoming common in shallower seas?

The discovery matters because it blurs the boundaries between the classic Ediacaran “chapters”. The Avalon and White Sea assemblages may not represent a clean handover, with one world disappearing and another suddenly replacing it.
Instead, the new Canadian fossils suggest overlap: Avalon-style frond-like organisms and more diverse White Sea-style animals may have shared the darkness and lived together in similar deep-water settings.
That makes early animal evolution look less like a sudden switch and more like a gradual ecological expansion. Animals were experimenting with new body shapes, new ways of living on the seafloor, and perhaps new ways of moving and feeding.
The roots of modern animal diversity may therefore lie in a long, uneven process that began in deeper marine environments far from the warmth of the Ediacaran sun, and before many animal groups became common in shallower seas.
A broader evolutionary idea
The study also raises a broader evolutionary idea.
Environments help shape life. A soft-bodied animal living on a quiet, deeper seafloor faced different challenges from one living in shallow water affected by waves, light, currents and shifting sediment. Those pressures can influence which body shapes and behaviours are useful and are passed on.
This is where the idea of convergent evolution can become helpful. Convergent evolution is when unrelated organisms evolve similar solutions to similar problems like wings in birds, bats and insects, or streamlined bodies in fish, dolphins and extinct marine reptiles.
In this sense evolution is repeated problem-solving under changing environmental rules over billions of years.
The same broad solutions, tubes, fronds, flattened bodies, may have been tried repeatedly as early animals explored the seafloor.
Over deep time, life can look uncannily inventive. But it’s shaped by the relentless testing ground of Earth itself.
Chris Kirkland, Professor of Geochronology, Curtin University and Anthony Clarke, Research Fellow, School of Earth and Planetary Sciences, Curtin University
This article is republished from The Conversation under a Creative Commons license. Read the original article.






So, once again, what creationists would like to pretend is a problem for evolution turns out to be nothing of the sort. The discovery of Ediacaran fossils earlier, more widely distributed, and in deeper-water settings than previously recognised does not contradict the evolutionary account; it enriches it. It adds detail to a picture that was already being assembled from geology, palaeontology, comparative biology and evolutionary theory.
The important point is that these fossils are not in the wrong sequence. They are not mammals in the Precambrian, birds before fish, or flowering plants before land plants. They are Ediacaran organisms in Ediacaran rocks, exactly where evolution says such organisms should be found. The adjustment is to the fine detail of their timing, distribution and ecology, not to the broad fact of life’s long evolutionary history.
Creationism, by contrast, has no useful explanation for any of this. It does not predict the Avalon, White Sea and Nama assemblages; it does not predict the orderly succession of fossils through deep time; it does not predict why soft-bodied Ediacaran organisms should be found in rocks hundreds of millions of years older than any modern mammal, bird or flowering plant. Its only response is to deny, distort or dismiss the evidence.
Science does something very different. It treats new evidence as information, not as a threat. If fossils are found earlier than expected, scientists ask why. If they are found in a different environment, scientists revise their understanding of the ecology. If they are found on a different palaeocontinent, scientists refine their view of how widely these organisms had spread. That is not weakness; it is the strength of a method that is accountable to reality.
And that is the lesson creationists never seem able to learn. Evolutionary biology can cope with new facts because it is a scientific explanation, not a sacred conclusion. Creationism cannot cope with them because its conclusion was fixed before the evidence was ever examined. The fossils from the Mackenzie Mountains therefore do not merely illuminate the strange world of the Ediacaran seas; they also illuminate the gulf between science, which follows the evidence, and dogma, which tries to make the evidence disappear.
Advertisement












All titles available in paperback, hardcover, ebook for Kindle and audio format.
Prices correct at time of publication. for current prices.

{kind=link}
{kind=link}
No comments :
Post a Comment
Obscene, threatening or obnoxious messages, preaching, abuse and spam will be removed, as will anything by known Internet trolls and stalkers, by known sock-puppet accounts and anything not connected with the post,
A claim made without evidence can be dismissed without evidence. Remember: your opinion is not an established fact unless corroborated.