

The experimental plants pollinated by bumblebees displayed significant differences on limestone soil (r) and tuff soil (l).
Image: UZH.
A clutch of papers published today serve to refute creationism by showing how evolution occurs in response to environmental selectors with no need to include magic or supernatural deities in the explanation. They also serve to highlight the lie that creationist cult leaders fool their dupes with that mainstream biologists are turning to creationism because the Theory of Evolution is inadequate for explaining the observable evidence.
On the contrary, these papers illustrate the robust nature of the TOE in explaining how plants and fish evolve in response to natural selectors, and the power of the theory to explain the observable evidence.
The first two are from substantially the same team led by biologist Florian Schiestl of the Department of Systematic and Evolutionary Botany at the University of Zurich, Switzerland, and deal with the way flowering plants respond to insects in their environment; both as beneficial pollinators and as harmful herbivores.
How has the interactions between flowering plants and insects, both pollinators and herbivores, affected the evolution of the plants? The interactions between flowering plants (angiosperms) and insects, including both pollinators and herbivores, have significantly shaped the evolution of flowering plants. These interactions have led to a variety of adaptations and co-evolutionary dynamics that have influenced the diversification, morphology, and reproductive strategies of flowering plants. Here’s a detailed look at how these interactions have impacted plant evolution:The researchers have shown that there is a complex relationship between the plants, the soil they grown on and the presence or absence of bumblebees and aphids. The soil-type may be an important element in the equation because the plants need different nutrients to enable them to adapt.
Pollinators and Plant Evolution
- Diversification and Speciation:
- Pollinator Specialization: Flowering plants often evolve specialized traits to attract specific pollinators. This can lead to reproductive isolation and speciation. For example, plants that are pollinated by bees may develop different floral structures, scents, and colors compared to those pollinated by birds or bats.
- Co-evolution: Plants and their pollinators engage in a co-evolutionary "arms race," where changes in one species drive changes in the other. For instance, the length of a flower’s nectar spur may evolve in tandem with the length of a pollinator’s proboscis.
- Morphological Adaptations:
- Flower Structure: The shape, size, color, and scent of flowers are adapted to attract specific pollinators. For example, flowers pollinated by moths tend to be white or pale (visible in low light) and have strong fragrances, whereas bird-pollinated flowers are often red and tubular.
- Nectar and Pollen: The production of nectar and the accessibility of pollen are tailored to the feeding habits of pollinators. This ensures efficient pollination while minimizing wastage.
- Reproductive Strategies:
- Temporal Synchrony: Flowering times may be synchronized with the activity periods of key pollinators. This temporal adaptation ensures that plants are pollinated when their pollinators are most active.
- Reproductive Assurance: Some plants evolve mechanisms like self-compatibility to ensure reproduction in the absence of reliable pollinators.
Herbivores and Plant Evolution
- Defensive Adaptations:
- Physical Defenses: Plants may develop structural defenses such as thorns, spines, and tough leaves to deter herbivores.
- Chemical Defenses: The production of secondary metabolites (e.g., alkaloids, tannins, and terpenoids) can reduce herbivory by making plants toxic, unpalatable, or harder to digest.
- Induced Defenses: Some plants can produce chemicals in response to herbivore attack, enhancing their defenses when needed.
- Mutualistic Relationships:
- Ant-Plant Interactions: Some plants form mutualistic relationships with ants, where ants protect the plants from herbivores in exchange for food or shelter.
- Predator Attraction: Plants may release volatile organic compounds (VOCs) that attract predators or parasitoids of herbivores, thus reducing herbivore pressure.
- Growth and Reproductive Strategies:
- Compensatory Growth: Plants may evolve the ability to regrow tissues rapidly after herbivore damage, reducing the impact of herbivory on fitness.
- Reproductive Allocation: Plants may allocate more resources to reproduction when under herbivore attack to ensure that they can produce offspring even under stress.
Combined Effects of Pollinators and Herbivores
- Trade-offs and Balance:
- Resource Allocation: Plants must balance resource allocation between attracting pollinators and defending against herbivores. This trade-off can shape the evolution of plant traits.
- Floral Scent and Herbivores: Some floral scents that attract pollinators may also attract herbivores. Plants may evolve complex scent profiles to balance these interactions.
- Indirect Effects:
- Herbivore-Induced Changes: Herbivory can indirectly affect pollination by changing plant traits. For example, herbivore damage to leaves can alter floral traits and reduce pollinator visits.
- Pollinator Behavior: The presence of herbivores can influence pollinator behavior and preferences, further affecting plant reproduction and selection pressures.
In summary, the interactions between flowering plants and insects, both pollinators and herbivores, have driven significant evolutionary changes in plants. These interactions have led to a diversity of adaptations that enhance reproductive success, deter herbivory, and optimize resource allocation. The co-evolutionary dynamics between plants and insects continue to shape the biodiversity and ecological complexity of plant-insect interactions.
How they carried out these experiment is explained in a news release from Zurich University:
Interaction with Insects Accelerates Plant EvolutionThe first of these open access papers explains the relationship between soil type and divergent evolution when the pollinator is a bumblebee:
A team of researchers at the University of Zurich has discovered that plants benefit from a greater variety of interactions with pollinators and herbivores. Plants that are pollinated by insects and have to defend themselves against herbivores have evolved to be better adapted to different types of soil.
Plants obtain nutrients and water from the soil. Since different soil types differ in their chemical and physical composition, plants need to adapt their physiology to optimize this process on different soil types. This evolutionary process leads to the formation of ecotypes, i.e., locally adapted “plant breeds” that differ slightly in appearance and may no longer be easily crossbred. The latter effect is considered to be the first step toward the formation of separate species. The adaptation of crops to local soil types is also crucial for agricultural productivity.
Experiment with bumblebees and aphids
A team of researchers led by biologist Florian Schiestl of the Department of Systematic and Evolutionary Botany at the University of Zurich has now discovered that the interaction of plants with pollinators and herbivorous insects influences their adaptation to soil types and thus the formation of ecotypes. In a two-year experiment, about 800 swede plants were grown over 10 generations on different soil types in a greenhouse. One group was pollinated by bumblebees, another by hand; in addition, the plants were cultivated with and without aphids (as herbivores).
At the end of the evolutionary experiment, the researchers investigated the extent to which the plants on the two soil types differed in shape and composition and how well they had adapted to the soil. In terms of shape, it was found that only the plants pollinated by bumblebees showed clear differences between the soil types, while the hand-pollinated plant groups remained largely the same.
Plants pollinated by bumblebees adapt best
When it comes to adaptation to soil types, the researchers even found significant adaptation only in bumblebee-pollinated plants with aphids after the two years of experimental evolution, while no significant adaptation to soil types was observed in the other groups.
The study, which was funded by the Swiss National Science Foundation (SNSF), also identified several genes that may play a critical role in this adaptation process. The results show that biotic interactions can have a strong influence on plants’ ability to adapt to abiotic factors and that adaptation is most efficient when plants are exposed to a variety of interactions.
Literature:
Dorey, Thomas and Schiestl, Florian P. Bee-pollination promotes rapid divergent evolution in plants growing in different soils. Nature Communications. 27 March 2024. DOI: 10.1038/s41467-024-46841-4
Dorey, T., Frachon, L., Rieseberg, L.H., Kreiner, J., Schiestl, F.P. Biotic interactions promote local adaptation to soil in plants. Nature Communications. 18 June 2024. DOI: 10.1038/s41467-024-49383-x
AbstractIn summary, what this research showed was that different soil types drove evolutionary diversification, probably because different nutrients were needed for the plants to diversify, and that biotic pollination as opposed to hand pollination enabled the plants to respond to the different plant types.
Divergent evolution leads to variation among populations and thus promotes diversification. In plants, adaptation to different soils, pollinator guilds, and herbivores is thought to be a key ecological driver of adaptive divergence, but few studies have investigated this process experimentally. Here we use experimental evolution with fast cycling Brassica rapa plants to study the impact of soil, pollination, herbivory, and their interactions on divergent evolution in various traits during eight generations of selection. We found significant evolutionary changes in plant phenotypes caused by all three factors and their interactions. In the richer soil type, plants showed higher evolutionary rates, especially with bumblebee-pollination, which led to the evolution of increased attractiveness of plants to bumblebees. Plants that had experienced aphid-herbivory showed lower attractiveness. We found the strongest evolutionary divergence when plants evolved in different soils with bee-pollination rather than hand-pollination, irrespective of herbivory. This “soil-pollinator effect” impacted divergence in diverse suites of traits, for example leaf size, flowering time, flower petal length, some floral volatiles and leaf glucosinolates. We conclude that the interaction between soil and biotic pollination may be an important cause for divergent evolution of plants growing on different soil types, even without a shift in pollinator guilds.
Introduction
The first step in plant speciation is the establishment of genotypic and phenotypic differences among populations, sometimes called plant races1. Although both neutral (i.e. genetic drift) and adaptive evolutionary processes can cause such variations, adaptation driven by selection is thought to be of prime importance for divergent evolution2. Factors that cause adaptation to local conditions include abiotic factors such as climate3, mixed abiotic-biotic factors like soil and soil-born microorganisms4, as well as biotic interactions such as those between plants, herbivores, and pollinators5,6,7. Whereas shifts in physiological-, root- and overall growth-related traits are thought to be driven primarily by soil adaptation8, divergence in defense mechanisms is expected to be selected for by variable herbivore communities9, and floral divergence is thought to be driven by shifts in pollinator species or pollinator guild composition10,11.
Divergent evolution in response to soil-type is common in plants and is well documented12. Plant adaptations to soil may be driven by physical and/or chemical parameters such as water-holding capacity, nutrient content13,14, salinity15,16, calcium ions in calcareous soil14,17, or heavy metal content in serpentine-18 and other metalliferous soils19,20. Soil represents the major source of nutrients for plants21,22, and selection favors mechanisms that improve resource acquisition and/or utilization23. Adaptation to physical properties typically causes alterations of the root system, and re-allocations between above- and below ground biomass24. On the other hand, physiological mechanisms to cope with chemical properties like the exclusion or tolerance of high concentration of toxic compounds in plant tissue have also been demonstrated12,25.
Plants in their natural environment are involved in multiple interspecific biotic interactions, for example with animals that pollinate flowers and/or feed on plant tissue26,27. By feeding on plants, herbivores induce strong selection on defense mechanisms in plants that may differ depending on the feeding mode and sensitivity of herbivore taxa to defense compounds9,28,29. Pollinators, in contrast, act mutualistically and select for signals and morphological traits ensuring optimal pollen transfer26,30. As pollinators differ in both their sensory preferences and morphological features, selection imposed by different pollinators can lead to divergent evolution among plant populations/species31,32.
Multiple lines of evidence show that different ecological factors such as soil-type and plant-animal interactions influence each other at various levels. First, soil-type strongly impacts the morphology and physiology of plants through phenotypic plasticity, and thereby changes the plants’ interactions with pollinators and herbivores33. For example, soil nutrients have been shown to impact plant phenology, flower production, flower shape, nectar production, as well as nectar and pollen chemistry34,35,36, all with high relevance for pollinator behavior. Plants growing on serpentine soils have been shown to receive less pollinator visitations and less herbivory37, whereas fertile soils can increase plant attractiveness to herbivores38. Hyperaccumulation of toxic metals is known to negatively affect herbivores and may also have an impact on pollinators and plant-fertility related traits39,40,41. Such environmentally-induced changes in plant traits likely change patterns of phenotypic selection caused by the behavior of pollinators and herbivores33,42, leading to evolutionary changes driven by interactions between soil and animal interactors.
In addition, although different ecological factors may typically select for different plant traits, the evolution of those traits is often not independent because of ecological and/or genetic mechanisms43,44,45,46, potentially resulting in trade-offs for plants47. For instance, soil chemistry can modify adaptive responses to both plant-herbivore and plant-pollinator interactions through resource limitation. Soil fertility can determine the evolution of plant growth and defense as predicted by the resource availability hypothesis48,49. In poor growing conditions, plants are expected to evolve low growth-rates but high levels of constitutive defense, a strategy driven by the lack of resources necessary to compensate for herbivore damage or produce effective herbivore-induced defenses48,50,51,52,53. In contrast, where resource availability is higher, plants may invest less in defense but evolve an ability to compensate for or tolerate herbivore damage54,55,56. In addition, traits that evolve in response to herbivores are intricately linked with those that evolve under selection by pollinators and vice versa57. For example, evolution of defense or herbivore-induced plasticity can compromise plant attractiveness to pollinators by decreasing the attractiveness of floral signals58,59, and rewards60,61, thereby causing a trade-off between pollinator attractiveness and defense against herbivores.
Finally, patterns of macroevolutionary diversification suggest combined effects of soil-type and pollinators on plant speciation in some plant lineages and floristic provinces8. Evidence for this is found in closely related plant species being adapted to different pollinators as well as to different soil types62,63,64,65. The evolution of such combined adaptions can be explained by adaptation to one factor, e.g. soil-type, driving the subsequent adaptation to the other (e.g. pollinator), either because pollinator distribution co-vary with soil-type in a mosaic-like fashion (i.e. the Grant-Stebbins model of pollinator driven speciation6,10), or soil-adaptation reinforces differential pollinator attraction upon secondary contact63. Both scenarios assume a shift in pollinator guilds as the primary force for divergence in flower-types and the subsequent evolution of pollination ecotypes32,66. Other studies, however, suggest that pollination ecotypes can also arise as a byproduct of habitat adaptation, such as to elevation ranges67,68,69. Generally, however, we know little about the combined effects of various ecological factors, such as soil, pollinators and/or herbivores in adaptive diversification, and their importance in promoting divergent evolution at the onset of the diversification process70.
A powerful way to study the process of diversification is experimental evolution71, which has, however, up to now only rarely been applied to plants, and most studies that do, focus on the effects of only one ecological factor31,61,72,73,74. Here we aim to contribute to filling this gap by disentangling the effects of soil, bee-pollination, and aphid-herbivory in shaping divergent evolution, by performing an eight-generation evolution experiment with fast cycling Brassica rapa plants as an outcrossing model with short generation time and high standing genetic variation75,76. In order to apply as-natural-as-possible ecological parameters in the experiment, we mimicked natural conditions found in populations of a Southern Italian Brassica species, Brassica incana, that grows on limestone- or tuff soil, the latter with higher nutrient availability than the former, with bumblebees and aphids being major biotic interactors. Nevertheless, the objective of the study was not to investigate evolutionary patterns specific to any plant species, but to provide overall insights and create new testable hypotheses about how soil and animal interactors impact plant divergent evolution. We used a 3-way factorial design with two different soil types (limestone- and tuff soil) that were collected in natural populations of Brassica incana, with- and without aphid-herbivory, and with either bumblebee-pollination or hand-pollination (Fig. 1). Hand pollination was included as a control for pollinator-mediated selection, as it represents random pollination without pollinator-mediated selection. Each treatment comprised two replicates with 49 plants each, evolving independently during eight consecutive generations; to reduce maternal effects, two generations without insects were included before plants were phenotyped. The soil used in the experiment was not sterilized, because sterilized soil can hardly be kept sterile during the experiment, with unnatural re-establishment of microbes being the possible consequence. We assessed the evolutionary responses of plants after the experiment by phenotyping plants in a common garden environment with standardized soil. We measured morphological as well as chemical traits, namely flower volatiles collected by headspace sorption and analyzed by gas chromatography, and leaf glucosinolates analyzed by liquid chromatography. These different traits are traditionally thought to evolve in response to different ecological factors such as pollinators (floral scent, flower morphology), herbivores (defense chemistry, i.e. glucosinolates), and soil (vegetative traits such as leaves). We performed various statistical analyzes, such as principle component analysis, linear discriminant function analysis and general linear models, and calculated evolutionary rates in haldanes. We addressed the following questions: (1) How do plant morphology, nectar, floral scent, defense traits, and attractiveness to pollinators evolve in response to different soil types, pollination and herbivory treatments? (2) Do interactions between these factors lead to distinct evolutionary trajectories and which combinations lead to the strongest divergence? For (1) we predicted that low nutrient soil should limit evolutionary response to bee-pollination and aphid-herbivory, as resource limitation is expected to induce strong trade-offs between traits. For (2) we predicted that because the ecological factors likely selected for different traits, more factors should cause more divergence77, and thus the interaction of both herbivory and pollination lead to the strongest divergence among plant populations on different soil types.
Our results show rapid divergent evolution in many traits driven by soil, pollination, herbivory and their interactions in our model system. We document that divergent evolution in response to different soil types was more pronounced with bee-pollination rather than hand pollination. This shows that pollination, the biotic factor with the strongest fitness-impact in our experiment, interacted with soil differences leading to the evolution of different plant phenotypes without a switch in pollinator quantity or quality. We conclude that divergent evolution driven by different soils can be accelerated by biotic pollination, which may act as a starting point for soil-pollinator driven diversification in plants.
Fig. 1: Design of our experimental evolution study.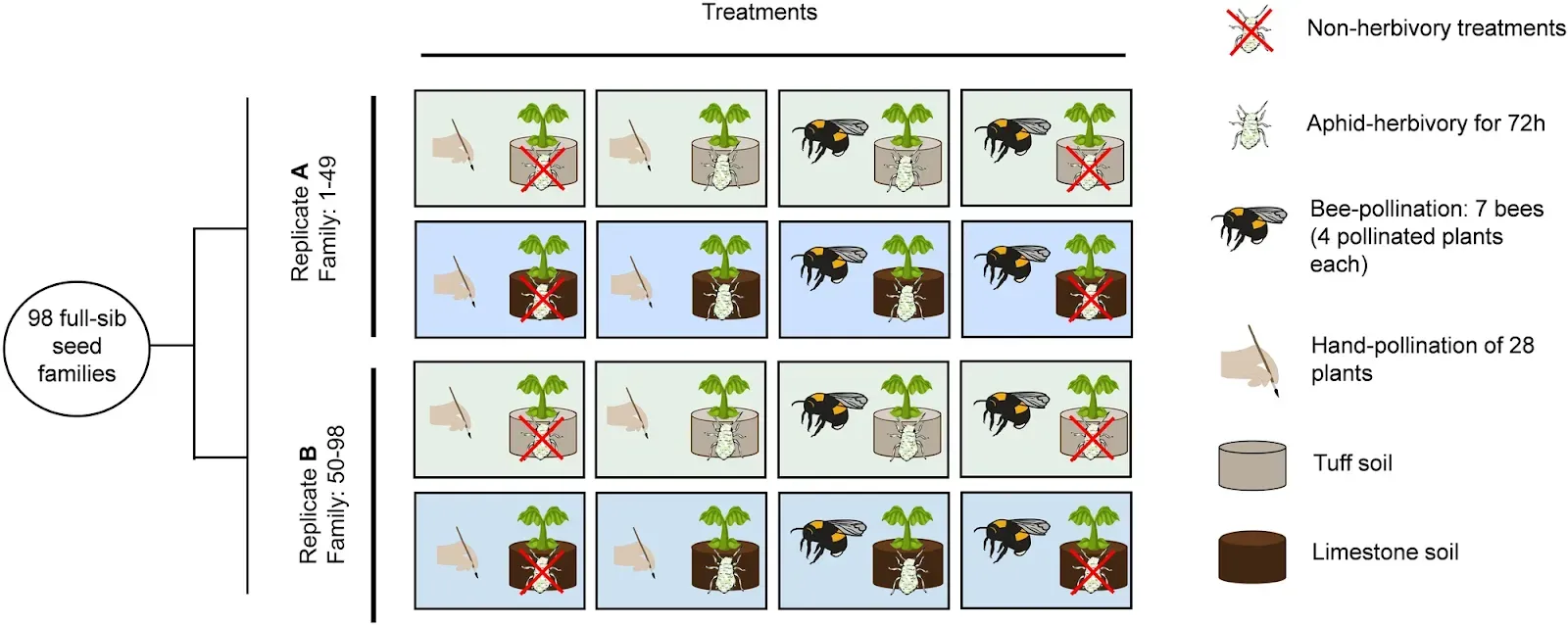 98 full-sib seed families of fast cycling Brassica rapa plants were divided into two replicates (A and B), and spread among treatment groups, with each seed family being represented in each treatment (49 plants were used per treatment per replicate). Differently colored squares represent the two types of soil, tuff- (pale green) and limestone soil (pale blue).Dorey, T., Schiestl, F.P.
98 full-sib seed families of fast cycling Brassica rapa plants were divided into two replicates (A and B), and spread among treatment groups, with each seed family being represented in each treatment (49 plants were used per treatment per replicate). Differently colored squares represent the two types of soil, tuff- (pale green) and limestone soil (pale blue).Dorey, T., Schiestl, F.P.
Bee-pollination promotes rapid divergent evolution in plants growing in different soils.
Nat Commun 15, 2703 (2024). https://doi.org/10.1038/s41467-024-46841-4
Copyright: © 2024 The authors.
Published by Springer Nature Ltd. Open access.
Reprinted under a Creative Commons Attribution 4.0 International license (CC BY 4.0)

The next paper deals with the effects of both pollinators and herbivory as plants adapt to changes in soil type.
AbstractIn summary, these experiments show a complex interaction between how the plants respond to mutualist insects in the form of pollinating bumblebees and antagonist herbivore insects in the form of aphids and how this facilitates adaptation to soil types.
Although different ecological factors shape adaptative evolution in natural habitats, we know little about how their interactions impact local adaptation. Here we used eight generations of experimental evolution with outcrossing Brassica rapa plants as a model system, in eight treatment groups that varied in soil type, herbivory (with/without aphids), and pollination mode (hand- or bumblebee-pollination), to study how biotic interactions affect local adaptation to soil. First, we show that several plant traits evolved in response to biotic interactions in a soil-specific way. Second, using a reciprocal transplant experiment, we demonstrate that significant local adaptation to soil-type evolved in the “number of open flowers”, a trait used as a fitness proxy, but only in plants that evolved with herbivory and bee pollination. Whole genome re-sequencing of experimental lines revealed that biotic interactions caused a 10-fold increase in the number of SNPs across the genome with significant allele frequency change, and that alleles with opposite allele frequency change in different soil types (antagonistic pleiotropy) were most common in plants with an evolutionary history of herbivory and bee pollination. Our results demonstrate that the interaction with mutualists and antagonists can facilitate local adaptation to soil type through antagonistic pleiotropy.
Introduction
Adaptation is a key process in evolution, leading to the emergence and modification of traits and the macroevolutionary diversification of organisms1.1. Adaptation at the population level is typically associated with evolutionary genetic changes that optimize the performance of organisms in their specific habitat, a phenomenon called local adaptation2.1,3.1. Local adaptation can be due to conditional neutrality, in which alleles are adaptive in one habitat, but neutral in the other, and/or antagonistic pleiotropy (i.e., a genetic trade-off), where different alleles at a locus are favored in different environments4.1,5.1,6.1,7.1. The relative importance of these mechanisms in local adaptation remains unclear8.1,9.1. Antagonistic pleiotropy is considered critical for maintaining genetic variation among natural populations, but direct experimental proof of this mechanism remains scarce6.1,7.1,8.1,10.1,11.1,12.1. To detect the genomic basis of local adaptation, most studies use reciprocal transplant experiments and measure fitness-marker associations in organisms in the local versus a foreign environment13.1. The detection of genetic trade-off is, however, hampered by the fact that fitness-marker associations must be significant in two environments and that temporal variation in environmental conditions may mask such associations during a particular season7.1. Therefore, long-term field studies or experimental evolution approaches provide a more powerful approach to detect antagonistically pleiotropic loci14.1. For example, in an 8-year field study on Arabidopsis thaliana conducted in Italy and Sweden, four fitness QTLs displayed a pattern of antagonistic pleiotropy, and seven showed that of conditional neutrality8.1. Other studies have used herbarium genomics or resurrection experiments to detect antagonistic selection in loci across populations, leading to strong population differentiation for certain loci15.1,16.1. Also, the genomic basis of local adaptation, whether it typically involves few versus many genes, and their relation to phenotypic traits is poorly understood17.1. Besides maintaining genetic diversity, local adaptation may also promote ecological speciation, through the emergence of ecotypes and eventually isolating barriers between differently adapted populations18.1,19.1.
Local adaptation in plants occurs through shifts in phenotypic and phenological traits and has been detected in response to climate variation16.1,20.1,21.1 or soil, driven by chemical factors such as salinity22.1, carbonate23.1, or heavy metal content in serpentine soils24.1. The role of biotic factors in local adaptation, such as mycorrhizal mutualists25.1, herbivory26.1 or pollination27.1 is much less well understood28.1,29.1. Plants can locally adapt to biotic interactors, if such interactions are spatially heterogeneously distributed across the landscape, in a geographic mosaic-like fashion. In plants, such local adaptation to the pollinator- or herbivore “climate” (sensu30.1) has been shown previously and is thought to be the basis for population variation in floral traits and defense mechanisms27.1,31.1. Besides driving local adaptation directly, biotic factors may also interact with abiotic factors and thus modify the selection strength and adaptive response of plant populations in an indirect way29.1. For example, the availability of soil nutrients may modify trade-offs between defense against herbivores and the ability of a plant to re-grow any lost tissue (i.e., tolerance). For this reason, herbivory may primarily select for resistance in nutrient-poor soils, whereas in nutrient-rich soils, it may favor tolerance (i.e., re-growth of lost tissue). This concept is called the growth-defense trade-off32.1,33.1. Despite the importance of multiple ecological factors for local adaptation, few studies consider more than one factor and biotic interactions such as herbivory or animal pollination are often not included29.1,34.1. Hence, we lack even a basic understanding of how multiple ecological factors interact to shape local adaptation35.1. In this study, we tested the hypothesis that biotic interactions, by changing patterns of selection and evolutionary trade-offs, impact the local adaptation of plants to soil type.
Soil is a key ecological factor for plants, as it forms a main source of nutrients and water, and enables interactions between plants and soil microbiomes36.1. Many studies have shown that plants adapt to different soil types, through physiological and morphological mechanisms, leading to the formation of soil ecotypes37.1. Sometimes, these soil ecotypes co-vary with floral- and defense traits suggesting that adaptation to soil types can be linked to adaptation to biotic interactions (i.e., pollinators and herbivores)32.1,38.1. We thus focused our investigation on local adaptation to soil type and the indirect effects of biotic interactions to this kind of adaptation. We focused on the following specific questions: 1) How fast does local adaptation evolve? 2) Is local adaptation to soil type impacted by selection imposed through herbivory and/or bee-pollination? 3) Is local adaptation in our system primarily driven by antagonistic pleiotropy or conditional neutrality at the genomic level?
We used experimental evolution with fast cycling Brassica rapa plants and a 3-way factorial design with two different soil types (limestone- and tuff soil, collected in nature and not sterilized), with- and without aphid herbivory, and with either bumblebee- or hand-pollination (i.e., eight treatment groups, Supplementary Fig. 1). Each treatment was replicated two times with 49 plants in each replicate evolving independently during eight consecutive generations, leading to a total number of 784 plants per generation. Eight generations with selection were followed by two generations without insects (i.e., hand-pollination) to reduce maternal effects caused especially by aphid-herbivores. First, to assess phenotypic evolution, plants of generation one and ten of all treatment groups were grown in the soil type they had evolved in, in the greenhouse. Plants were only grown in their “local” soil for this analysis to avoid any soil-induced plastic effects and thus allow for a (within soil-type) comparison of trait evolution. We quantified several plant traits to gain an as-comprehensive-as-possible view of the phenotypic changes that may have evolved.
Second, we conducted a reciprocal transplant experiment, where plants of generation ten of all treatment groups were grown in the soil they had evolved in (local soil), as well as the other soil type (foreign soil). To assess evolutionary changes, plants of generation one were re-grown on both soils along with plants from the tenth generation (i.e., as a resurrection experiment). Altogether in this experiment, 1376 plants were grown and phenotyped, of which 1118 randomly chosen individuals were subsequently genotyped by whole-genome resequencing. In addition, bioassays were performed with bumblebees in flight cages to assess the attractiveness of 912 randomly selected plants using choice assays. All phenotyping and bioassays were done without aphids being present on the plants. To assess local adaptation on the phenotypic level, we chose the trait “the number of open flowers” at pollination day, which had the strongest and most consistent positive association with “relative seed set” and “bumblebee first choices”, as assessed in data from generation one and two in this experiment (Supplementary Tables 1,2). Seed set could not be assessed directly in our local adaptation experiment, because the DNA sampling after the bioassays left many of the smaller plants with a significant proportion of their biomass removed. For the “local” vs “foreign” criteria for local adaptation to be true3.1, we expected that plants that evolved in either soil type would always outperform plants that were transplanted into this soil type. Statistically, this would be shown by a significant soil line x soil ( = G x E) interaction and a significant post hoc linear contrast between the “local” (e.g., limestone lines in limestone soil) and “foreign” (e.g., tuff lines in limestone soil) treatments. “Soil lines” were the plant genotypes having evolved in a particular soil, and “soil” was the soil type that plants were grown in during the transplant experiment (i.e., the environment).
In a previous study on phenotypic selection, using plants of generations one and two of this experiment39.1, we showed that pollinator-mediated selection varies according to soil type and the presence/absence of herbivory, despite only one type of pollinator being used in this experiment. Selection diverged the most between plants grown in different soils, with herbivory and bee pollination. Based on these different figures of selection, we hypothesized that the strongest local adaptation would evolve in treatment groups with biotic interactions where the stronger divergent selection was observed. Because more fertile soil allows for a more rapid evolutionary response40.1, we also expected stronger patterns of local adaptation in tuff soil.
Here we show that local adaptation to different soil types evolved only in plants that interacted with herbivores and bee pollinators. Whole genome re-sequencing of experimental plants revealed that biotic interactions led to a tenfold increase in SNP markers with significant allele frequency change and that plants that had interacted with herbivores and bee-pollinators showed the most markers with opposite allele frequency change in the different soil types (antagonistic pleiotropy). We conclude that biotic interactions speed up the evolution of local adaptation to soil types in plants and that antagonistic pleiotropy is a key mechanism for driving this evolutionary process.Fig. 1: Trait evolution in the evolution experiment. The figure shows the difference in phenotype (mean ± s.e.m. values) among treatments in plants of generation ten when growing in “local” soil (gray bars), and generation one (black bars; plants of generation one and ten were grown together). Differences in phenotype were estimated by two-sided linear mixed models (LMM). To discriminate evolutionary changes from changes due to soil-induced plasticity, we considered only plants grown in their local soil in this analysis. a Plants’ attractiveness to bumblebees, (b) number of open flowers on the day of pollination, and (c) flower diameter. Plants with an evolutionary history of bumblebee pollination were more attractive to bumblebees (a) consistent with earlier studies47.1,57.1, had more open flowers (b) and produced larger flowers (c) in tuff soil than plants that evolved with hand-pollination. Plants in limestone soil that evolved with herbivory had more open flowers than those without herbivory (b). Asterisks indicate statistically significant effects of soil, herbivory, and pollination (P < 0.05; further statistical values are shown in Supplementary Tables 3–4). The dashed lines distinguish plants growing in limestone (left side) from those growing in tuff (right side). Attractiveness of plants was tested separately for soil types and herbivory groups, because of the strong differences in attractiveness between them (plants in tuff soil and those that evolved without herbivory were generally the most attractive). Attractiveness (N = 189 visits for 456 plants) and trait values (N = 443 limestones, 456 tuff) were assessed without aphids on the plants (i.e., without the effects of herbivore-induced plasticity).
The figure shows the difference in phenotype (mean ± s.e.m. values) among treatments in plants of generation ten when growing in “local” soil (gray bars), and generation one (black bars; plants of generation one and ten were grown together). Differences in phenotype were estimated by two-sided linear mixed models (LMM). To discriminate evolutionary changes from changes due to soil-induced plasticity, we considered only plants grown in their local soil in this analysis. a Plants’ attractiveness to bumblebees, (b) number of open flowers on the day of pollination, and (c) flower diameter. Plants with an evolutionary history of bumblebee pollination were more attractive to bumblebees (a) consistent with earlier studies47.1,57.1, had more open flowers (b) and produced larger flowers (c) in tuff soil than plants that evolved with hand-pollination. Plants in limestone soil that evolved with herbivory had more open flowers than those without herbivory (b). Asterisks indicate statistically significant effects of soil, herbivory, and pollination (P < 0.05; further statistical values are shown in Supplementary Tables 3–4). The dashed lines distinguish plants growing in limestone (left side) from those growing in tuff (right side). Attractiveness of plants was tested separately for soil types and herbivory groups, because of the strong differences in attractiveness between them (plants in tuff soil and those that evolved without herbivory were generally the most attractive). Attractiveness (N = 189 visits for 456 plants) and trait values (N = 443 limestones, 456 tuff) were assessed without aphids on the plants (i.e., without the effects of herbivore-induced plasticity).

Together, these two papers show how the interaction between multiple environmental factors drives evolution and how an organism doesn't exist in isolation but is very much the product of its environment, resulting in a superficial appearance of design for living in that environment. Natural selection, without any magical interference, by reducing the frequency of detrimental mutations and favouring the survival of beneficial mutations makes adaptation and the appearance of design, inevitable.
Creationists, with their teleological thinking defect retained from childhood find it impossible to understand how this all happens without intelligent agency telling organisms how to react - which of course assumes the organisms such as these brassica plants are also sentient and capable of following instructions to change their allele frequencies over time.
What Makes You So Special? From The Big Bang To You

How did you come to be here, now? This books takes you from the Big Bang to the evolution of modern humans and the history of human cultures, showing that science is an adventure of discovery and a source of limitless wonder, giving us richer and more rewarding appreciation of the phenomenal privilege of merely being alive and able to begin to understand it all.
Available in Hardcover, Paperback or ebook for Kindle
Ten Reasons To Lose Faith: And Why You Are Better Off Without It

This book explains why faith is a fallacy and serves no useful purpose other than providing an excuse for pretending to know things that are unknown. It also explains how losing faith liberates former sufferers from fear, delusion and the control of others, freeing them to see the world in a different light, to recognise the injustices that religions cause and to accept people for who they are, not which group they happened to be born in. A society based on atheist, Humanist principles would be a less divided, more inclusive, more peaceful society and one more appreciative of the one opportunity that life gives us to enjoy and wonder at the world we live in.
Available in Hardcover, Paperback or ebook for Kindle

No comments :
Post a Comment
Obscene, threatening or obnoxious messages, preaching, abuse and spam will be removed, as will anything by known Internet trolls and stalkers, by known sock-puppet accounts and anything not connected with the post,
A claim made without evidence can be dismissed without evidence. Remember: your opinion is not an established fact unless corroborated.