
In the face of overwhelming evidence contradicting the creationist view of reality, it’s difficult to avoid the conclusion that such beliefs are rooted in a deep-seated psychological need—one that drives wilful ignorance and a denial of the observable world.
Consider, for example, the mountain of data showing that Earth is not a few thousand years old, nor was life magically created in its present form within days. Instead, the planet is over four billion years old, and the diversity of life we see today is the product of a long, gradual evolutionary process, shaped by environmental change and punctuated by countless extinctions.
One small yet compelling piece of that vast evolutionary puzzle comes in the form of an exquisitely preserved, 444-million-year-old fossil, discovered some 25 years ago in a South African quarry. Only recently has it been identified as a stem-group arthropod by Professor Sarah Gabbott of the University of Leicester. She outlines her findings in an article for The Conversation and a detailed research paper published in *Papers in Palaeontology.
Professor Gabbott's article in The Conversation is reprinted here under a Creative Commons license, reformatted for stylistic consistency:
Marine fossil found in South Africa is one of a kind, thanks to unusual preservation
Sarah Gabbott, University of Leicester
A fossilised creature found in a South African roadside quarry 25 years ago has finally got an official name. The small, segmented, crustacean-like creature, dated to 444 million years ago, can now be introduced as Keurbos susanae. It belongs to the arthropod group of animals, which accounts for about 84% of all known species that exist today, including insects, spiders and crabs.
Palaeontologist Sarah Gabbott explains what’s so unusual about her discovery, which she named as part of the process of describing it scientifically.
What can you tell us about this creature and the environment it lived in?
The fossil is about 50cm long and has 46 almost identical segments. Projecting from each is a delicate, gill-like structure. It would probably have looked like a bit like a horseshoe crab and the gills would have been for absorbing oxygen from the water it lived in. Its insides are exquisitely well-preserved, which is very unusual for fossils – normally only the hard, more decay-resistant external features would be preserved. You can see bundles of muscle fibres that would have powered the limbs, tendons and an internal scaffold structure that gave the animal rigidity.
We think it would have spent most of its life living on, or more likely just above, the seafloor, probably walking and swimming in an undulatory (waving) motion.
It lived in the immediate aftermath of the end Ordovician extinction event more than 440 million years ago, caused by glaciations (the spread of icy conditions) across vast swaths of the planet. This extinction wiped out about 85% of Earth’s species. The marine basin that Keurbos susanae inhabited was probably very cold and at times covered with sea ice.
It was a relatively hostile environment in other ways too. Our analyses of the chemistry of the shales – the sediments on the sea bed where this animal and others lived, now turned to rock – shows that they were deposited under anoxic conditions (that is, there was no oxygen circulating freely in the water). And at times free hydrogen sulfide occurred in the sediment porewaters (the water in tiny spaces between grains of sediment) and even above the seafloor. Not much could live in these conditions and this was critical to this fossil’s amazing preservation.
It meant the carcass was not scavenged by other animals after it died. Also, the chemistry was important in the process whereby the soft tissues, which should usually rot away rapidly, became mineralised quickly after death. This turned the animal’s anatomy to mineral which survived for hundreds of millions of years until it was discovered.
It is preserved “inside out”.
Keurbos susanae is a new genus and species which we are still trying to place among other early arthropods. The fact that its insides are better preserved than its outside makes it difficult to compare with other fossils that are preserved the “other way round”.
How did you find the fossil and what else has been found in that area?
The site is in the Cedarberg mountains, north of Cape Town. To collect fossils in this area you need a permit granted by the Council for Geoscience. Fossil-bearing rocks are protected by law because of their heritage and scientific value.
Fossil hunting in these rocks takes a lot of hard work and patience, splitting open the shales with a hammer and chisel. These shale rocks are what’s left of layers of silt that were once on the sea floor. The fossils here are super rare: you can dig and split shale for days and not find a single fossil! But we know there are some in there because of discoveries made previously.
I found two specimens. The first one is complete but the second one only has the middle part of the body preserved.
In the same rocks we have found some of the earliest vertebrate fossils with mineralised teeth, called conodonts. They were eel shaped and predatory. Also eurypterids (sea scorpions), arthropods with powerful swimming appendages, which would have cruised through the frigid waters. There are also orthocones – a type of chambered cephalopod – like the mollusc fossils called ammonites, which have been found in large numbers, but with a straight shell instead of coiled.
Why has it taken 25 years to describe Keurbos susanae scientifically?
Two reasons really.
First, because of the nature of preservation, where all the insides are perfectly preserved but the outside (the carapace or body covering) is absent, it is just difficult to interpret and compare to other fossils. And secondly because the specimen’s head and legs are missing and these are key characteristics that palaeontologists would use to help them to understand the evolutionary relationships of such fossils.
If more specimens were to be found, with their heads and legs, we could be more certain about where this fossil fitted in the scheme of life. But the site where I found it has been covered in a lot of rock from quarrying activity. So we decided to describe what we had in the meantime, and not wait for more examples.
The fossil’s name, Keurbos susanae, refers to the place where I found it and to my mother, Sue, who encouraged me to follow a career that made me happy, whatever that might be.
Sarah Gabbott, Professor of Palaeontology, University of Leicester
This article is republished from The Conversation under a Creative Commons license. Read the original article.
The identification of Keurbos susanae as a stem-group arthropod offers yet another clear window into the deep history of life on Earth—an ancient organism exquisitely preserved from a world 444 million years ago. It stands as one more piece of evidence among countless others supporting the reality of evolution and the immense age of our planet. Yet, as so often happens, creationists will either ignore this discovery or move the goalposts—demanding evidence not as a genuine request for understanding, but as a rhetorical tactic to appear reasonable while remaining impervious to whatever is presented.Abstract
A new exceptionally preserved euarthropod, Keurbos susanae gen. et sp. nov. from the Upper Ordovician Soom Shale Konservat-Lagerstätte of South Africa, is described herein. Two specimens exhibit an unusual preservation style such that the cuticular exoskeleton is preserved in low relief but retains high-fidelity details, whereas the internal anatomy is preserved in three dimensions and includes myoanatomy and aspects of the connective inter- and intrasegmental endoskeleton. The trunk has 46 homonomous segments and tapers towards the posterior. The dorsal surface comprises the posterior margins of tergites that form rounded paratergal folds at their lateral margins. Sternal morphology is seen from an internal view of the fossils and includes axial sternites and associated ovoid plates, both with fringing setae along their posterior margins. Patchy, incomplete preservation of the appendages is consistent with the ‘reversed’ taphonomy of these fossils in which sclerotized elements that project beyond the body margin, such as lamellae with vascular channels and pits, are preserved, whereas those inside the body margins are absent or poorly expressed. Euarthropod characters include an arthrodized tergal and sternal exoskeleton, and a segmental connective endoskeleton. Recent depopulation of the euarthropod stem group has witnessed conspicuously arthrodized fossils formerly placed in the stem group being shifted into the crown, increasing the likelihood that Keurbos is likewise a crown-group euarthropod. Comparison with a phylogenetically disparate suite of similarly homonomous, multisegmented taxa indicates no convincing synapomorphies. The grouping of relevant middle Palaeozoic exemplars as ‘enantiopod’ pancrustaceans could suggest that Keurbos might be allied.
Euarthropods (chelicerates, myriapods and pancrustaceans including the insects) are a well-represented and diverse phylum in the fossil record. The earliest euarthropod body fossils date to c. 521 Ma (Daley et al. 2018) and there is a coherent picture of their subsequent radiation and evolution captured in exceptionally preserved fossil deposits (Edgecombe & Legg 2014; Edgecombe 2020; Aria 2022).
Much of our knowledge of the anatomy of early Palaeozoic arthropods is derived from sites of Burgess Shale-type preservation. These biotas provide a record of decay-resistant extracellular structures such as the cuticular exoskeleton and appendages, as well as more labile/decay-prone cellular tissues such as the gut and musculature (Butterfield 2002, 2003; Vannier et al. 2014.1; Strang et al. 2016; Young & Vinther 2017). In addition, the same deposits can record details of the neural anatomy in euarthropods (e.g. Ma et al. 2012; Cong et al. 2014.2; Ortega-Hernández et al. 2022.1).
Burgess Shale-type preservation presents two broad taphonomic pathways: the first, the most frequently occurring, is that in which more recalcitrant external structures are preserved as essentially two-dimensional carbonaceous compressions; the second, much rarer pathway is that in which labile internal structures (guts and muscles) are captured in three-dimensional relief, through early authigenic mineralization, most typically by calcium phosphate. In cases in which silica preserves musculature this is thought to be secondary replacement of an earlier phosphate phase (Young & Vinther 2017). The Late Ordovician fossils described in this contribution present a unique taphonomic style that shares some aspects with Burgess Shale-type preservation but is also distinct. Except for anatomy that projects beyond the body margin the external cuticular features are either not preserved or occur as low-relief imprints. The majority of fossil anatomy is represented by three-dimensionally preserved internal anatomy. As such the fossils provide a view of the endoskeleton and myoanatomy that is known in few other fossil euarthropod taxa.
The purposes of this contribution are: (1) to taxonomically describe a new genus and species from the Soom Shale Konservat-Lagerstätte and place constraints on its position in the euarthropod tree; and (2) to describe its internal anatomy and endoskeleton and compare this with other examples in the Palaeozoic, and to provide a taphonomic basis for anatomical interpretations.
GEOLOGICAL SETTING
The Soom Shale Member is the basal member of the Cedarberg Formation, which lies directly above a glaciogenic diamict, the Pakhuis Formation; it outcrops in the Western Cape Province of South Africa. The Soom Shale hosts exceptionally preserved fossils of latest Hirnantian to earliest Rhuddanian age; based on the chitinozoan biota a Late Ordovician age is most likely (Vandenbroucke et al. 2009). Both the age of the shales and their sedimentology indicate deposition at the end of the Ordovician glaciation event in a shallow marine basin (Gabbott et al. 2010, 2017.1). Fine-grained distal turbidites intercalate with organic-rich layers consisting of marine snow and wind-blown loess grains. Bounding the Soom Shale there are sub-areal glacial pavements in the underlying Pakhuis Formation, and evidence of shallow-water marine sediments in the overlying Disa Siltstone Member of the Cedarberg Formation. The glacially influenced setting of the Soom Shale is unusual for a Konservat-Lagerstätte: at its base dropstones demonstrate sea-ice cover, and most of the sediment was sourced from a landscape recently exposed by retreating ice (Young & Nesbitt 1998).
The Soom Shale is parallel-laminated, black when unweathered and presents geochemical signatures (high total carbon, high molybdenum) indicative of anoxic and at times euxinic conditions in the pore and bottom waters. Endobenthic and benthic taxa are absent or extremely rare. Despite this, several nektobenthic taxa, such as eurypterids, naraoiids, lobopodians and trilobites, suggest that life could inhabit, perhaps briefly, the bottom waters of the basin. Most taxa are nektonic and include conodonts, orthoconic cephalopods, caryocarids, undescribed anapsid-like fish and possible medusae (for details see Gabbott et al. 2017.1 and references therein).
The taphonomy of the fossils in this deposit is unusual. Many exhibit details of non-biomineralized anatomy as illitic clays and/or as organic films with rare pyrite coatings. Alunite group minerals, formed later during weathering of the shales, are also associated with soft tissues but played no role in their preservation (Gabbott 1998.1; Gabbott et al. 2001). Thus, in general Soom Shale fossils have some preservation modes in common with the spectrum of Burgess Shale-type preservation that includes, principally, carbonaceous films with subsidiary mineralization in clays, pyrite and calcium phosphate (Butterfield 1990; Gabbott et al. 2004; Page et al. 2008; Farrell 2014.3; Anderson et al. 2021a). In the Soom Shale, originally biomineralized anatomy, including calcium phosphate, calcite and aragonite, is mouldic, having undergone extensive dissolution. This occurred during diagenesis because pore waters of very low pH are indicated by geochemical redox-sensitive proxies (Gabbott 1998.1, Gabbott et al. 2001).
FIG. 1
Holotype C1002, Keurbos susanae gen. et sp. nov. A, complete specimen of the part. B, line drawing showing the main morphological features. To aid orientation the exoskeletal anatomy is shaded: pink, sternites; orange, ovoid plates. Abbreviations: lvl, lower vascularized lamella; uvl, upper vascularized lamella. Scale bar represents 20 mm.FIG. 2
Paratype C2044, Keurbos susanae gen. et sp. nov. A, the specimen part. B, line drawing showing the main morphological features. To aid orientation the exoskeletal anatomy is shaded: blue, tergite boundaries; pink, sternites; orange, ovoid plates. Abbreviations: ptf, paratergal fold; uvl, upper vascularized lamella. Scale bar represents 20 mm.FIG. 3
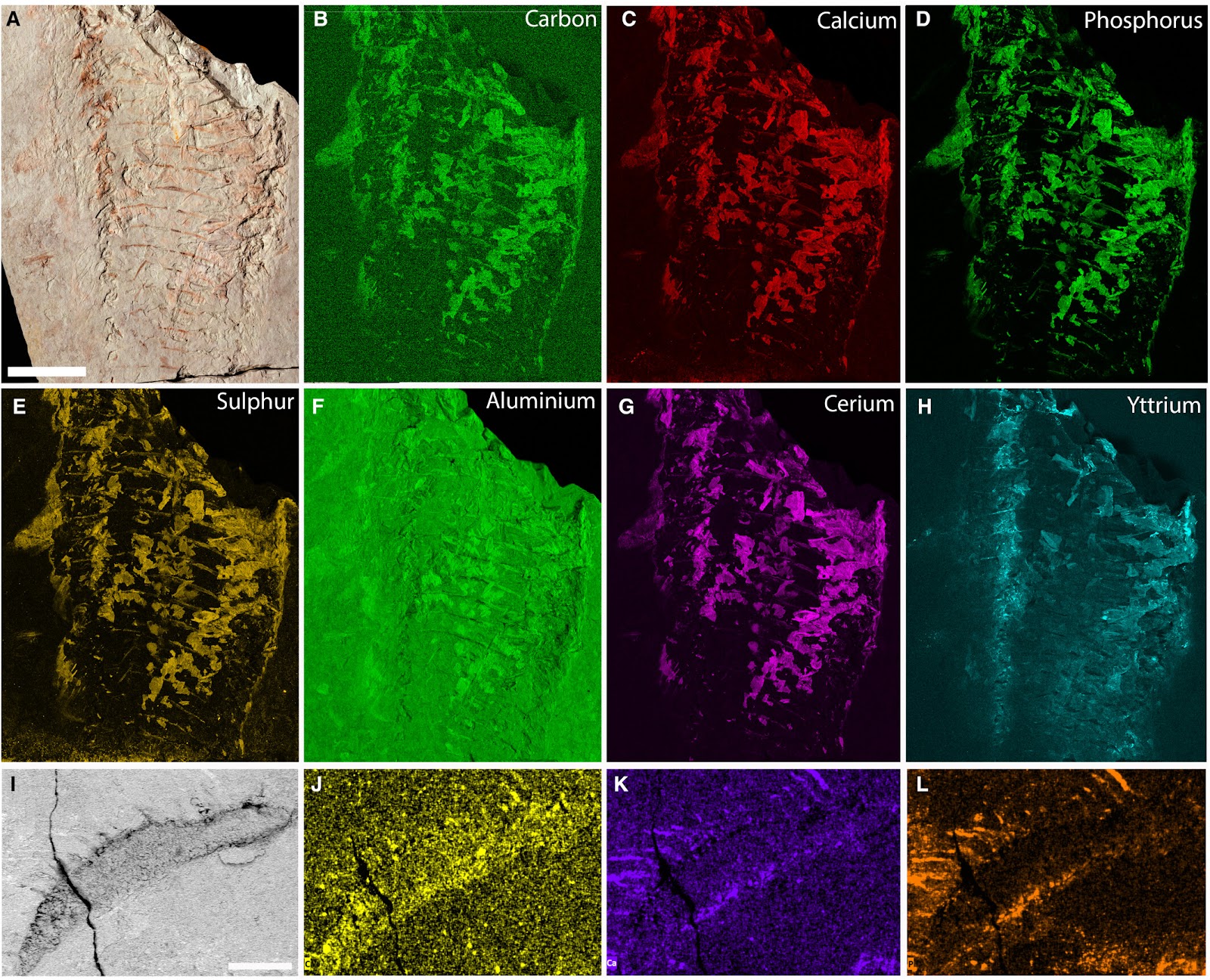
Taphonomy of Keurbos susanae gen. et sp. nov. A–H, paratype C2044. I–L holotype C1002. In all images lighter areas indicate relatively higher abundance of the elements. A, paratype counterpart specimen photograph of the anterior block; B–H, μXRF analysis of the specimen. I, back-scattered electron image of anterior setose structure; J–L, EDX maps: J, carbon map; K, calcium map; L, phosphorus. Scale bars represent: 20 mm (A–H); 500 μm (I–L).FIG. 4
Keurbos susanae gen. et sp. nov. Anatomy of the cephalon in the holotype C1002. A, counterpart, showing bilaterally symmetrical disposition of the cephalic anatomy that tapers anteriorly (above the horizontal crack) in relation to the trunk; dashed rectangle indicates the location of C. B, arcuate setose structure on the right side in A. C, muscle fibres and arcuate setose structure on the right side in A. D, composite image showing the anatomy of the cephalic region on the counterpart; white arrowhead indicates subcircular patches of robust material with a granular texture; black arrowhead indicates a narrow linear feature. Abbreviations: as, arcuate setose structure; m, muscle. Scale bars represent: 10 mm (A, D); 1 mm (B, C).FIG. 5
Dorsal morphology of Keurbos susanae gen. et sp. nov. A–C, paratype C2044; D–G, holotype C1002. A, counterpart; white arrowheads indicate tergites that are broken (see also Figs 3, 11). B, part showing three tergites; white arrowhead shows position of C. C, close-up of regular ridges on the posterior margin of a tergite. D, impressions of dimples in the light-coloured sediment that may represent the texture of the dorsal exoskeleton are subtle but best seen at the midpoint along the axis. E, possible preservation of finely ribbed tergal exoskeleton. F, counterpart showing two tergites; dashed rectangle indicates the location of G. G, counterpart showing regular ridges on the posterior margin of a tergite. Abbreviations: op, sternal oval plate; t, tergite; tds, transverse dorsal structure. Scale bars represent: 10 mm (A, D); 5 mm (E, F); 1 mm (B, C, G).FIG. 6
Paratype C2044, showing paratergal fold morphology. A, right lateral half of part showing the imbricating folds, one for each segment (see Fig. 2 for a line drawing and Fig. 11 for an outline of the paratergal folds); white arrowhead indicates Promissum pulchrum conodont apparatus (also on B); white dotted rectangle shows the location of B. B, close-up of the paratergal folds showing their feathery texture, with two thickened lines with prominent relief on the anterior edge of each fold (black arrowheads). C, back-scattered electron image of thickened line at the anterior edge of a paratergal fold (black arrowhead) and small patches of black, carbonaceous material representing the texture of the paratergal fold; the texture mimics the white dashed line and forms multiple arcs roughly parallel to the outer margin of the paratergal fold. D, back-scattered electron image of two thickened lines at the anterior edge of the paratergal fold (black arrowheads) and between them a series of evenly spaced lines that are perpendicular to the thickened lines; the bright areas in the image are composed of elevated Ca and P and are calcium phosphate. Abbreviations: ptf, paratergal fold; uvl, upper vascularized lamella. Scale bars represent: 10 mm (A); 5 mm (B); 500 μm (C, D).FIG. 7
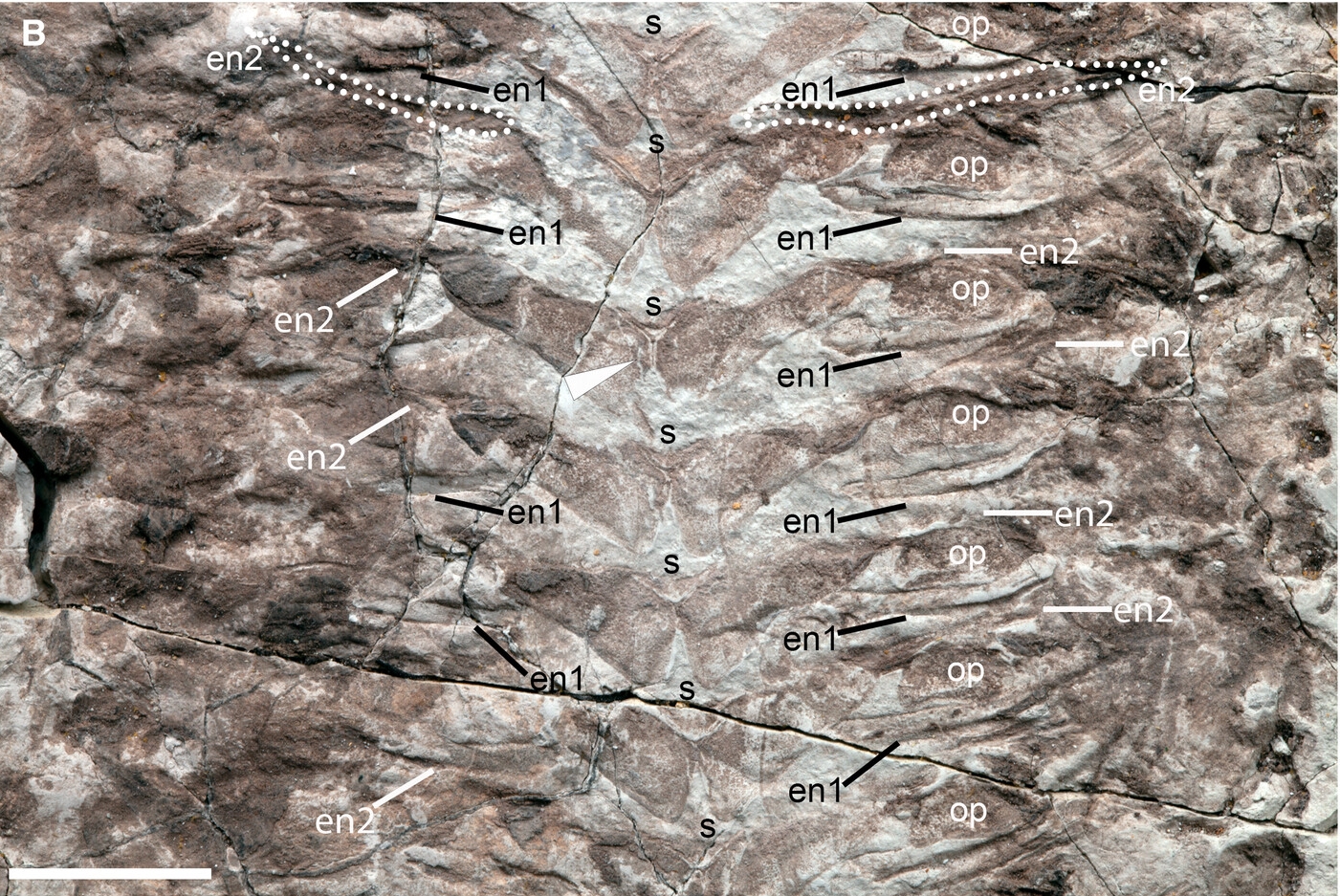
Ventral morphology of Keurbos susanae gen. et sp. nov. A–C, holotype C1002; D–F, paratype C2044. A, sternites, ovoid plates and endoskeleton; anteriorly the sternites are narrow (sag.) and do not meet at the midline (white sternite), in the posterior the sternites are broader and meet axially to form a Y-shaped structure (black sternite); the area between the white dashed and black dashed lines indicates the location of B and C, respectively. B, close-up of segments between white dashed lines in A, showing the axial Y-shaped structure where bilaterally symmetrical subrectangular sternites possess a thickened margin; there are two types of endoskeleton, en 1 representing possible tendons and en2 representing an extension of the sternite plate laterally, extending closer to the body margin (outlined in white dots). C, close-up of segments between black dashed lines in A showing, in addition to the morphology in B, the setae along the posterior margins of the sternites and oval plates (black arrowheads). D, sternites with three-dimensional thickened margins forming a Y-shaped structure at the axis, ornamented sternal ovoid plates (white dotted boundaries) and muscle fibres that overlie the ovoid plates. E, close-up of three of the ovoid plates outlined in D showing a characteristic spongy texture that is identical to the sternite texture (black arrowhead). F, three-dimensional relief of the sternite Y-shaped structure, which may be brittle and could break. Abbreviations: en1, endosternite 1; en2, endosternite 2; m, muscle; op, sternal ovoid plate; s, sternite; t, tergite. Scale bars represent: 10 mm (A–C); 2 mm (D–F).FIG. 8
Vascularized lamellae of Keurbos susanae gen. et sp. nov. (holotype C1002). A, section of the trunk with the best-preserved lamellae; the morphology of upper vascularized lamellae consists of a frayed and wrinkled lateral margin (black-outlined white arrowheads); a ridge along the midline (black arrowhead) from which extend posterolaterally a series of veins that form a gently scalloped posterior margin (white arrowheads); and a robust ‘strut’ that tapers to a point approximately two-thirds along the lamella margin (grey arrowheads). The white dashed rectangle indicates the area of B, and the black dashed rectangle indicates the area of C. B, fine details of the lower vascularized lamella; note faint veins extending posterolaterally from the central dark line (white arrowheads). C, two upper vascularized lamellae and underlying lower lamella (white arrowhead). Abbreviation: lvl, lower vascularized lamella. Scale bars represent: 20 mm (A); 5 mm (B, C); 1 mm (D).FIG. 9
Upper vascularized lamella of Keurbos susanae gen. et sp. nov. (holotype C1002). A, image of the counterpart of a vascularized lamella. B, camera lucida drawing of A showing anterior, robust strut (black arrowheads) and central ridge structure (grey arrowhead), and paratergal fold. Abbreviations: ptf, paratergal fold; v, vein. Scale bar represents 5 mm.FIG. 10
Keurbos susanae gen. et sp. nov. holotype C1002, counterpart pieces highlighting internal anatomy. A, pieces of the counterpart of the holotype with soft-tissue preservation of muscles and endoskeleton; muscle (brown material) is especially prevalent towards the lateral margins in the anterior and posterior of the trunk. B, internal anatomy and orientation of the muscle fibre bundles; dorsal arcuate structures, purple lines; endoskeletal type 1 rods (possible tendons), yellow dotted lines; endoskeletal type 2 rods, orange dotted lines; muscle fibres, white lines. Scale bar represents 20 mm.FIG. 11
Keurbos susanae gen. et sp. nov. paratype C2044, highlighting internal anatomy. A, section of the preserved trunk. B, internal anatomy and orientation of the muscle fibres indicated. The endoskeletal type 1 rods have variable morphological expression: appearing as an irregular ropey feature (white arrowheads) or as rod-like and flanked by a flange of tissue (black arrowheads). Endoskeletal type 1 rods (possible tendons), yellow dotted lines; muscle fibres, white lines; paratergal folds, white dashed lines; black-outlined white arrowhead, position of Promissum pulchrum conodont apparatus. Scale bar represents 10 mm.
Gabbott, S.E., Edgecombe, G.D., Theron, J.N. and Aldridge, R.J. (2025)
A new euarthropod from the Soom Shale (Ordovician) Konservat-Lagerstätte, South Africa, with exceptional preservation of the connective endoskeleton and myoanatomy. Pap Palaeontol, 11: e70004. https://doi.org/10.1002/spp2.70004
Copyright: © 2025 The authors.
Published by John Wiley & Sons, Ltd. on behalf of The Palaeontological Association. Open access.
Reprinted under a Creative Commons Attribution 4.0 International license (CC BY 4.0)










Their position is not shaped by evidence and never was. Science, by contrast, progresses precisely because it is shaped by evidence—questioning, testing, and revising in light of new discoveries like this one. Keurbos susanae is not just a scientific curiosity; it is a silent rebuttal to every claim that the fossil record lacks transitional forms or that complex life appeared fully formed. Reality, as always, speaks more clearly than denial ever can.
Advertisement
Amazon

Amazon

Amazon

Amazon

Amazon

Amazon

Amazon

Amazon

All titles available in paperback, hardcover, ebook for Kindle and audio format.
Prices correct at time of publication. for current prices.

No comments :
Post a Comment
Obscene, threatening or obnoxious messages, preaching, abuse and spam will be removed, as will anything by known Internet trolls and stalkers, by known sock-puppet accounts and anything not connected with the post,
A claim made without evidence can be dismissed without evidence. Remember: your opinion is not an established fact unless corroborated.