
fossil of Tametara mirim
Agustin Martinelli
80-million-year-old snake fossil sheds light on why lizards lost their limbs and started slithering
Of all the primitive ‘just-so’ stories in Genesis, the tale of how the serpent was condemned to crawl on its belly must rank among the most transparently mythological. Having supposedly tempted Eve to eat the forbidden fruit, God curses the serpent to go on its belly and ‘eat dust’ for the rest of its life. Although the story never explicitly says that the serpent previously had legs, the curse plainly implies a change and has traditionally been understood as an explanation for why snakes slither instead of walking.
Like so much of Genesis, it begins with an observation familiar to its authors—snakes have no legs—and invents a magical tale to explain it. It is an ancient example of an etiological myth: folklore devised to explain why something in the natural world appears as it does, long before anyone had the evidence or intellectual framework needed to understand its real history.
Even as mythology, however, the story is strangely incoherent. Genesis itself does not identify the serpent as Satan but introduces it as the most cunning of the wild animals God had made. Nevertheless, this supposedly ordinary animal can speak, reason and somehow knows about a prohibition that God had given to Adam before Eve had even been created. The text makes no attempt to explain how it acquired either this knowledge or the power of speech.
More awkward still for anyone insisting that the story is literally true, the serpent’s predictions prove substantially more accurate than God’s warning. God tells Adam that he will die on the day he eats the fruit. The serpent tells Eve that they will not die, that their eyes will be opened and that they will become like God, knowing good and evil. They eat the fruit, their eyes are opened and they do not die that day. God then confirms that the humans have indeed become ‘like one of us’ in knowing good and evil. According to Genesis itself, Adam goes on to live for another 930 years.
On the most straightforward reading of the story, therefore, the serpent tells Eve the truth while God’s threat to Adam proves false. Later theologians have attempted to rescue the tale by inventing a ‘spiritual death’, or by claiming that Adam merely began to die that day, but neither qualification appears in the text. These are apologetic repairs added by people who can see the problem but cannot admit that the story’s authors created it.
Science, meanwhile, has uncovered a very different account of how snakes acquired their extraordinary bodies—one involving neither talking reptiles nor fruit, curses or human disobedience, but descent with modification over immense periods of geological time. Snakes are highly modified members of the squamate reptile radiation. Their ancestors progressively elongated their bodies, increased the number of vertebrae and reduced their limbs as different lineages adapted to particular ways of life. Their evolutionary history extends back at least to the Middle Jurassic, about 160 million years ago.
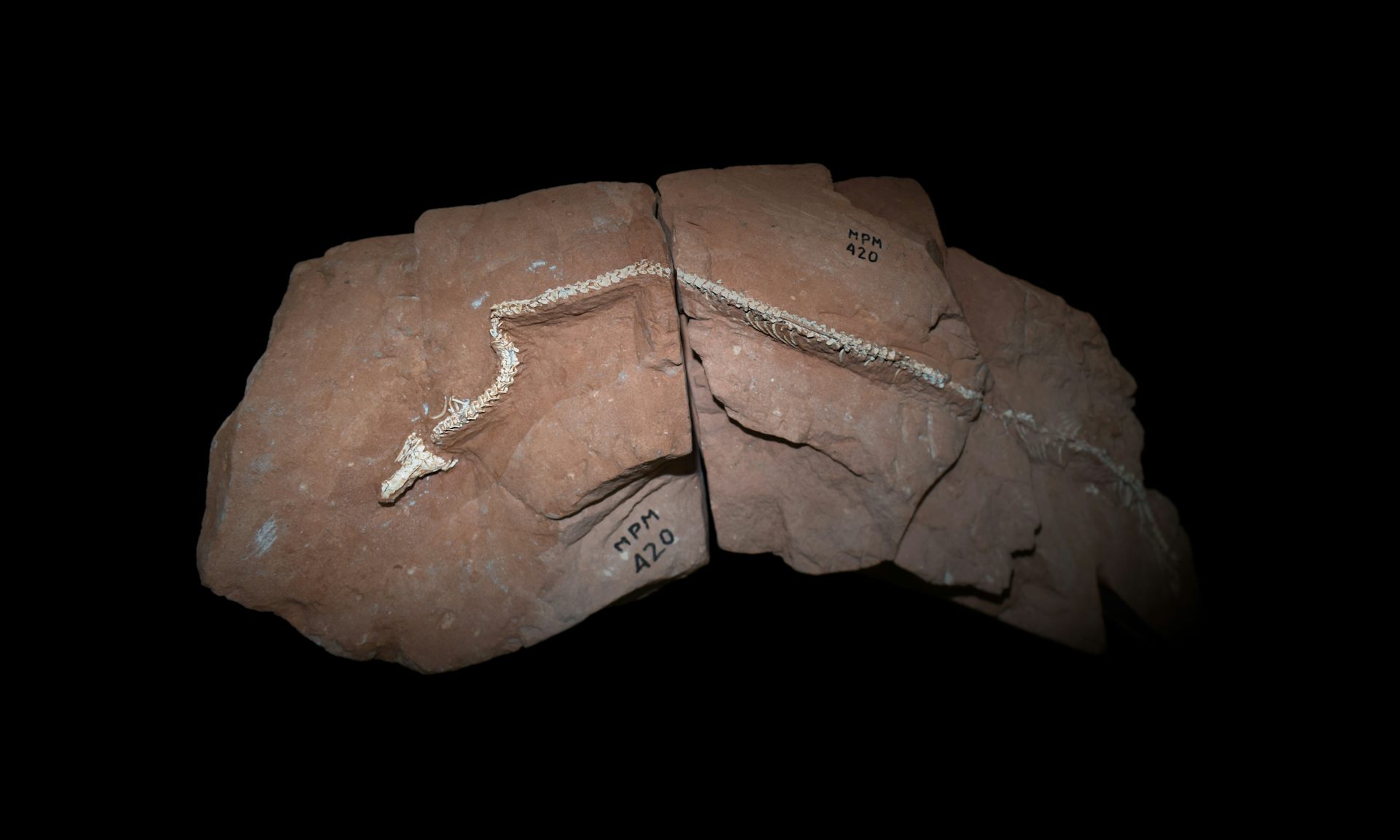
Now, a paper published in Nature by an international team led by Tiago R. Simões describes an exceptionally well-preserved early snake from the Late Cretaceous of Brazil. Named Tametara mirim, the animal lived approximately 85–75 million years ago, when non-avian dinosaurs still dominated terrestrial ecosystems. It is the first articulated snake fossil known from Brazil and preserves a three-dimensional skull together with more than 100 connected vertebrae.
The preserved portion of its skeleton appears to have lacked forelimbs. Because the region around the pelvis is missing, the researchers cannot determine whether it retained vestigial hindlimbs, as some other early snakes did. Its skull also combines recognisably snake-like specialisations with ancestral features resembling those of other squamate reptiles—precisely the sort of mosaic of old and new characteristics expected in an evolving lineage, but not from the instantaneous transformation of an animal by a divine curse.






























{kind=link}