
Holotype of Helmetia expansa USNM 83952, dorsal view.
Jean-Bernard Caron and Sarah Losso.

Creationist apologetics often involve attributing phenomena that science cannot yet explain to divine intervention, relying on the assumption that gaps in scientific knowledge serve as evidence for the existence of a deity. This approach frequently employs the false dichotomy fallacy, suggesting that if science lacks an explanation, then "God did it" by default. Such reasoning does not establish the existence of a god capable of addressing these gaps; instead, it often depends on the cultural biases of the audience to support this conclusion.
However, science continually seeks to fill these gaps with evidence-based explanations. A recent example is the formal description of the Cambrian fossil Helmetia expansa, which provides insights into arthropod evolution during the Cambrian Period. This period is often mischaracterized by creationist narratives as a time when multiple distinct body plans appeared suddenly without ancestral forms, purportedly indicating a single act of creation.
The study, published in the *Journal of Systematic Palaeontology* by Professor Sarah R. Losso and Javier Ortega-Hernández of Harvard University's Museum of Comparative Zoology and Department of Organismic and Evolutionary Biology, along with Jean-Bernard Caron of the Royal Ontario Museum's Department of Natural History (Palaeobiology Section) in Toronto, Canada, offers a comprehensive analysis of Helmetia expansa. Their research reveals that Helmetia expansa is an arthropod related to trilobites, shedding light on the evolutionary diversification of this group during the Cambrian. This finding provides evidence of gradual evolutionary processes, countering claims of abrupt appearance without ancestral lineage.
The study is further explained in a press release from Harvard University, which details how this research resolves a 100-year-old mystery surrounding this Cambrian arthropod.
What information do you have on Helmetia expansa and their occurrence in the fossil record? Helmetia expansa is an extinct arthropod from the middle Cambrian period, approximately 508 million years ago. Its fossils have been primarily discovered in the Burgess Shale of Canada. Initially identified by palaeontologist Charles Doolittle Walcott in 1918, the species remained inadequately described for over a century.
Recent studies have provided a comprehensive analysis of Helmetia expansa, revealing that it belongs to a rare group of early arthropods called concilitergans, which are close relatives of trilobites. Unlike trilobites, concilitergans lacked calcified exoskeletons, making their fossilization rare and typically occurring only under exceptional conditions like those found in the Burgess Shale.
The anatomy of Helmetia expansa includes a broad, leaf-shaped exoskeleton divided into a trapezoidal cephalon (head shield), a thorax with six segments, and a triangular pygidium (tail shield) with five marginal spines. Notably, the cephalon features an anterior sclerite with two median eye-like structures and two stalked lateral eyes positioned beneath the head shield. citeturn0search5 Recent examinations have identified broad gills and walking legs in several specimens, indicating that Helmetia likely exhibited both walking and swimming behaviours similar to trilobites.
The fossil record of Helmetia expansa is limited, primarily due to the delicate nature of its exoskeleton, which posed challenges for fossilization. Nonetheless, the specimens recovered from the Burgess Shale provide valuable insights into the morphology and evolutionary relationships of early arthropods. Comparative studies with related species from other Cambrian deposits, such as the Chengjiang biota in China and the Sirius Passet in Greenland, have enhanced our understanding of concilitergan diversity and their evolutionary significance.
Ancient fossil sheds big light on evolution mystery: solving a 100-year arthropod mystery
For over a century, the Cambrian arthropod Helmetia expansa remained a mystery. Discovered by paleontologist Charles Doolittle Walcott in 1918, it was initially classified as a crustacean. Despite frequent mentions in research papers, this species has never been formally described, and only one specimen has ever been illustrated.
Now, in a new study published in the Journal of Systematic Palaeontology, Harvard researchers led by Sarah Losso, postdoctoral fellow in the Department of Organismic and Evolutionary Biology, formally describe Helmetia expansa, offering new insights into its anatomy, behavior and evolutionary relationships.
Helmetia expansa belongs to a rare group of early arthropods called concilitergans, close relatives of trilobites. Unlike trilobites, concilitergans lacked calcified exoskeletons, so their remains only fossilized under exceptional conditions—like those in the 508-million-year-old Burgess Shale of Canada, where even soft tissues like guts, legs, and gills were preserved.
Although more specimens were collected, only one individual of Helmetia expansa had been figured and no study examined additional material in detail to formally describe the species or clarify concilitergan evolution.
We need to study more than one specimen to see the species’ full range of morphology and preservation.
Professor Sarah R. Losso, lead author
Museum of Comparative Zoology and Department of Organismic and Evolutionary Biology
Harvard University, Cambridge, MA, USA.
The team examined 36 specimens, at the Smithsonian Institution and the Royal Ontario Museum, from the Cambrian Period of the Burgess Shale of Canada. They photographed the specimens, both wet and dry, using a polarizing filter that can better reveal subtle features and extinction phases, and compared them to related species found in the Chengjiang biota in China, and one from the early Cambrian Sirius Passet in Greenland.
Helmetia had a leaf-like exoskeleton, with some specimens preserving eyes, medial eyes, digestive systems and limbs. Early arthropods had limbs with a walking leg for locomotion and food capture, and a gill used for respiration. On the holotype, only the gills are visible – leading past researchers to believe Helmetia lacked legs and swam exclusively. But, the team found broad gills and walking legs in several specimens, showing it likely walked like trilobites.
Even more surprising were two specimens caught in the early stages of molting, a behavior never before documented in concilitergans.
Molting strategies have never been known in any concilitergan. All arthropods molt their hard exoskeletons to grow, but no one had seen this behavior before in a concilitergan because you have to catch a specimen in the act of molting, and it’s difficult to get just the right timing.
Professor Sarah R. Losso.
The molting specimens show the new exoskeleton closer to the edge of the head, suggesting the animal exited front the front of the body – similar to horseshoe crabs, which use an anterior exit strategy, unlike most crabs that exit from the rear of the body.
The researchers also discovered a wide range of adult body sizes as Helmetia grew. While the smallest specimen was only 92 millimeters long, one exceeded over 180 millimeters.
These patterns tell us not only how these 508-million-year-old animals grew, but how big they could get.
Professor Sarah R. Losso.
Based on updated interpretations of Helmetia expansa’s morphology, the researchers confirmed two main helmetiids groups: the Helmediidae (which includes Helmetia expansa) and is characterized by segment boundaries and side spines, and Tegopeltidae, which are marked by segment fusion and a lack of spines. The researchers also assigned Arthroaspis bergstroemi, known from Greenland since 2013, to the group Conciliterga.
Our findings give a much fuller picture of what Helmetia looked like, how it lived, and how concilitergans are related to each other,” Losso said, “which is very important for future studies on Conciliterga and other early arthropods.
Professor Sarah R. Losso.
AbstractDiscoveries such as the reclassification of Helmetia expansa exemplify how science continually addresses and closes the so-called “gaps” often cited by creationists—not only in the fossil record, but also in our broader understanding of evolutionary relationships among major taxonomic groups. In this case, the new insights into Helmetia's affinities reinforce the view that the Cambrian was not a singular, sudden explosion of fully formed, unrelated body plans, but a dynamic period of evolutionary experimentation and diversification. Stem-group species evolved, radiated, and in many cases, like the trilobites, eventually went extinct—following the same evolutionary mechanisms seen throughout life's history.
The trilobitomorphs are a megadiverse and ecologically versatile group of Paleozoic euarthropods that include the iconic trilobites, as well as non-biomineralized clades exclusively known from Konservat-Lagersttätten. The concilitergans, defined by the presence of a broad, variably effaced and flattened dorsal exoskeleton, have received comparatively little attention. This is particularly true for Helmetia expansa from the mid-Cambrian Burgess Shale (British Columbia) – the first chronologically reported species of Conciliterga, which was originally figured (based on a single specimen) by Walcott (1918). Here, we present the first comprehensive description of H. expansa based on all material available from the Burgess Shale, totalling 36 specimens housed at the Royal Ontario Museum and the Smithsonian Institution. The non-biomineralized dorsal exoskeleton is broad and flat, with a serrated margin throughout. The body consists of a cephalon with well-developed anterolateral spines, six thoracic tergites and a large pygidium bearing two pairs of lateral spines and a single terminal spine. The preserved appendages include short uniramous antennae followed by 15 pairs of homonomous and biramous limbs composed of a gracile endopodite and an exopodite with a broad lobe and thick lamellae. The digestive system consists of a straight gut tract, including a ‘J’-shaped foregut and five paired digestive glands on the anterior half of the body expressed as small and convex oblong structures with a submillimetric lamellar ultrastructure. Two specimens of H. expansa provide the first direct evidence of moulting in concilitergans. The lack of dorsal ecdysial sutures suggests a marginal moulting strategy similar to extant Xiphosura. A revised phylogeny of Trilobitomorpha supports Arthroaspis bergstroemi from the early Cambrian Sirius Passet of Greenland as the earliest branching concilitergan and prompts a new diagnosis for the clade. Our results inform the internal relationships within Conciliterga and formalize the families Helmetiidae (Helmetia, Rhombicalvaria, Haifengella and Kuamaia) and Tegopeltidae (Tegopelte, Skioldia, Saperion).
Introduction
Trilobitomorpha is a major clade within Artiopoda that includes diverse Paleozoic non-biomineralized euarthropods in addition to the iconic biomineralized trilobites (e.g. Berks et al., 2023; Edgecombe & Ramsköld, 1999; Lerosey-Aubril et al., 2017). The trilobitomorphs comprise various sub-groupings that are generally known from several Cambrian Konservat-Lagerstätten around the world, including well-defined clades such as nektaspids (e.g. Naraoia, Misszhouia; Mayers et al., 2019; Zhai et al., 2019.1), xandarellids (e.g. Xandarella, Sinoburius; Hou & Bergström, 1997; Chen et al., 2019.2) and concilitergans (e.g. Helmetia, Kuamaia; Hou & Bergström, 1997) (Fig. 1). In this context, concilitergans are of particular interest as they have been widely considered as either the sister group or at least close relatives of trilobites (Berks et al., 2023; Chen et al., 2019.2; Du et al., 2019.3; Edgecombe & Ramsköld, 1999; Mayers et al., 2019; Stein et al., 2013), although several analyses have otherwise recovered trilobites as sister to nektaspids (e.g. Legg et al., 2013.1; Lerosey-Aubril et al., 2017). Concilitergans are characterized by a broad and flattened dorsal exoskeleton, the variable effacement of the tergite boundaries in different species, and the presence of a prominent anterior sclerite accommodated in a cephalic notch in several taxa (Fig. 1). As traditionally defined, concilitergan diversity consists of eight Cambrian taxa (Table 1). However, the internal relationships within the group have never been specifically investigated. Beyond their original systematic descriptions, concilitergans as a clade have received less attention compared to nektaspids (e.g. Bond & Edgecombe, 2020; Budd, 1999.1; Mayers et al., 2019; Paterson et al., 2010; Pérez-Peris et al., 2020.1; Whittington, 1977; Zhai et al., 2019.4), xandarellids (e.g. Chen et al., 2019; Hou et al., 2019.5; Schmidt et al., 2021; X. Zhang et al., 2012) and even early branching trilobitomorphs or other artiopodans (e.g. Berks et al., 2023; Du et al., 2019.3; Schmidt et al., 2022; Stein & Selden, 2012.1; M. Zhang et al., 2022.1). Part of the complications for understanding concilitergan evolution is that their non-biomineralized body fossils are rare compared to other trilobitomorphs, with most described species to date being known from a limited number of figured specimens (e.g. Hou, 1987; Hou & Bergström, 1997; Whittington, 1985; Zhao et al., 2014).Concilitergans are chronologically among the earliest non-trilobite Cambrian euarthropods to be recognized from the Burgess Shale, with Helmetia expansa as the first recorded representative of this group, having been discovered in 1917 and informally named by Walcott (1918, p. 7) with a single illustrated specimen. That specimen was re-illustrated by Resser in 1931 in a posthumous publication (Walcott, 1931, p. 38, plate 23) and designated as the holotype, but no other material was mentioned, and no formal description was provided. H. expansa has not been described since then, and only the holotype specimen (USNM 83952) has been discussed in the context of other findings (Briggs, 1994; Edgecombe & Ramsköld, 1999; Ortega-Hernández, 2015; Simonetta & Delle Cave, 1975; Walcott, 1918). This is problematic, as H. expansa remains incompletely known and yet has been featured in numerous phylogenetic analyses that explore the evolution of Trilobitomorpha and Artiopoda more broadly (Berks et al., 2023; Chen et al., 2019; Edgecombe & Ramsköld, 1999; Hou & Bergström, 1997; Hou et al., 2019.4; Legg et al., 2013.1; Paterson et al., 2012; Schmidt et al., 2022.2; Stein et al., 2013). In this study, we formally describe Helmetia expansa based on the holotype and previously unstudied material at both the Royal Ontario Museum and the Smithsonian Institution, report new morphological data including evidence for moulting and a digestive system with paired glands and produce a new phylogeny for Conciliterga in the broader context of Trilobitomorpha.Table 1. Diversity and biostratigraphical distribution of Cambrian Conciliterga. Figure 1. Concilitergan diversity and relationships. A,Arthroaspis bergstroemi, MGUH 30382, from the early Cambrian (Stage 3) Sirius Passet, Greenland. B,Helmetia expansa, USNM 83952, from the mid-Cambrian (Wuliuan) Burgess Shale, Canada. C,Kuamaia lata, CJHMD 00064, from early Cambrian (Stage 3) Chengjiang, China. D,Tegopelte gigas, USNM 189201, from the mid-Cambrian (Wuliuan) Burgess Shale, Canada. E, simplified topology based on results from phylogenetic analyses herein which resolved Arthroaspis bergstroemi as an early-branching concilitergan.
Figure 1. Concilitergan diversity and relationships. A,Arthroaspis bergstroemi, MGUH 30382, from the early Cambrian (Stage 3) Sirius Passet, Greenland. B,Helmetia expansa, USNM 83952, from the mid-Cambrian (Wuliuan) Burgess Shale, Canada. C,Kuamaia lata, CJHMD 00064, from early Cambrian (Stage 3) Chengjiang, China. D,Tegopelte gigas, USNM 189201, from the mid-Cambrian (Wuliuan) Burgess Shale, Canada. E, simplified topology based on results from phylogenetic analyses herein which resolved Arthroaspis bergstroemi as an early-branching concilitergan.
 Figure 2. Holotype of Helmetia expansa USNM 83952, dorsal view. A, Cross polarized light. B, Low angle light. C, Magnification of A showing cephalon. D, Magnification of A showing lamellae. Abbreviations: cep, cephalon; anl, anterolateral spine; ant, antenna; asc, anterior sclerite; ey, eye; ex, exopodite; hyp, hypostome; lm, lamellae; mey, medial eyes; mg, midgut gland; opn, optical nerve; py, pygidium; rs, reniform structure; stn, sternite number; th, thorax.
Figure 2. Holotype of Helmetia expansa USNM 83952, dorsal view. A, Cross polarized light. B, Low angle light. C, Magnification of A showing cephalon. D, Magnification of A showing lamellae. Abbreviations: cep, cephalon; anl, anterolateral spine; ant, antenna; asc, anterior sclerite; ey, eye; ex, exopodite; hyp, hypostome; lm, lamellae; mey, medial eyes; mg, midgut gland; opn, optical nerve; py, pygidium; rs, reniform structure; stn, sternite number; th, thorax. Figure 3. Schematic drawing of Helmetia expansa holotype USNM 83952. Abbreviations: ant, antenna; ey, eye; ex, exopodite; hyp, hypostome; lm, lamellae; mey, medial eyes; mg, midgut gland; st, sternite.
Figure 3. Schematic drawing of Helmetia expansa holotype USNM 83952. Abbreviations: ant, antenna; ey, eye; ex, exopodite; hyp, hypostome; lm, lamellae; mey, medial eyes; mg, midgut gland; st, sternite. Figure 4. Helmetia expansa, ROMIP 66310, ventral view. A, cross polarized light. B, cross polarized light submerged in water. C, Magnification of box A1 showing gut with lumen infills and midgut glands. D, magnification of box A2 showing sternites. Abbreviations: an, anus; ant, antenna; att?, possible proximal limb attachment; ey, eye; ex, exopodite; g, gut tract; j, ‘J’-shaped foregut; m, moult; m als, moult of anterolateral spine; mg, midgut gland; stn, sternite number.
Figure 4. Helmetia expansa, ROMIP 66310, ventral view. A, cross polarized light. B, cross polarized light submerged in water. C, Magnification of box A1 showing gut with lumen infills and midgut glands. D, magnification of box A2 showing sternites. Abbreviations: an, anus; ant, antenna; att?, possible proximal limb attachment; ey, eye; ex, exopodite; g, gut tract; j, ‘J’-shaped foregut; m, moult; m als, moult of anterolateral spine; mg, midgut gland; stn, sternite number. Figure 5. Schematic drawing of Helmetia expansa ROMIP 66310. Abbreviations: an, anus; ant, antenna; ey, eye; ex, exopodite; j, ‘J’-shaped foregut; m, moult; m als, moult of anterolateral spine; mey, medial eyes; mg, midgut gland; rs, reniform structure; st, sternite.
Figure 5. Schematic drawing of Helmetia expansa ROMIP 66310. Abbreviations: an, anus; ant, antenna; ey, eye; ex, exopodite; j, ‘J’-shaped foregut; m, moult; m als, moult of anterolateral spine; mey, medial eyes; mg, midgut gland; rs, reniform structure; st, sternite.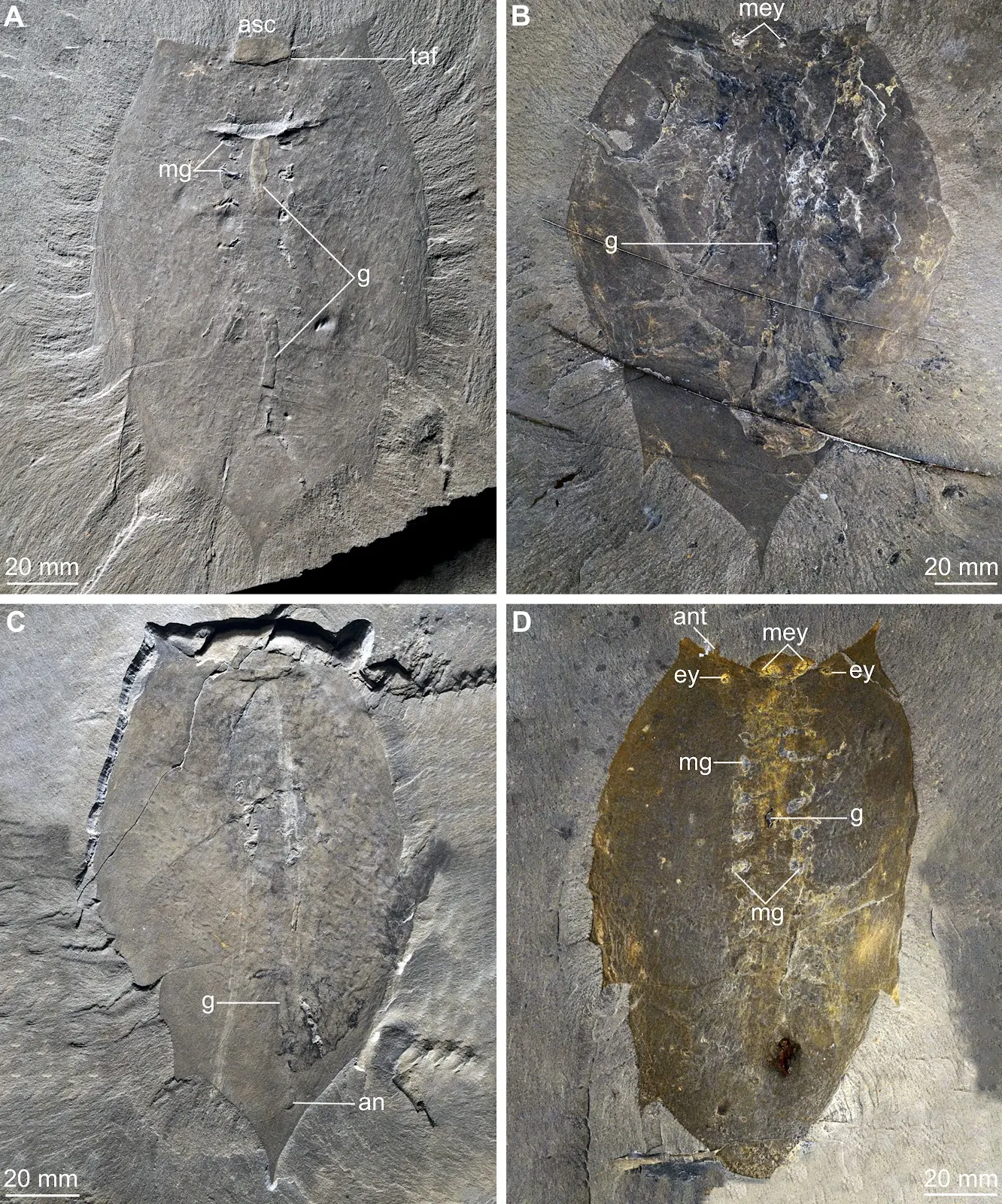 Figure 6. Complete specimens of Helmetia expansa. A, ROMIP 66311, ventral view. B, ROMIP 66303, dorsal view. C, ROMIP 66309, dorsal view. D, ROMIP 65115, dorsal view. Abbreviations: an, anus; ant, antenna; asc, anterior sclerite; ey, eye; g, gut tract; mey, medial eyes; mg, midgut gland; taf, transverse anterior fold.
Figure 6. Complete specimens of Helmetia expansa. A, ROMIP 66311, ventral view. B, ROMIP 66303, dorsal view. C, ROMIP 66309, dorsal view. D, ROMIP 65115, dorsal view. Abbreviations: an, anus; ant, antenna; asc, anterior sclerite; ey, eye; g, gut tract; mey, medial eyes; mg, midgut gland; taf, transverse anterior fold. Figure 7. Exopodites of Helmetia expansa. A, ROMIP 66308 ventral view. B, ROMIP 61038 dorsal view. C, USNM 213821 dorsal view. D, close up USNM 213821 box C1. E, ROMIP 66304 dorsal view. Abbreviations: ag, agnostid trilobite; ant, antenna; asc, anterior sclerite; ey, eye; ex, exopodite; mey, medial eyes; s, sternite.
Figure 7. Exopodites of Helmetia expansa. A, ROMIP 66308 ventral view. B, ROMIP 61038 dorsal view. C, USNM 213821 dorsal view. D, close up USNM 213821 box C1. E, ROMIP 66304 dorsal view. Abbreviations: ag, agnostid trilobite; ant, antenna; asc, anterior sclerite; ey, eye; ex, exopodite; mey, medial eyes; s, sternite. Figure 8. Endopodites of Helmetia expansa. A, USNM 189226 dorsal view. B, ROMIP 66313 dorsal view. C, magnification of pygidium in USNM 189226 showing endopodites. D, magnification of posterior thorax showing appendages. Abbreviations: asc, anterior sclerite; ey, eye; en, endopodite; hyp, hypostome; lm, lamellae; pb, podomere boundary; taf, transverse anterior fold.
Figure 8. Endopodites of Helmetia expansa. A, USNM 189226 dorsal view. B, ROMIP 66313 dorsal view. C, magnification of pygidium in USNM 189226 showing endopodites. D, magnification of posterior thorax showing appendages. Abbreviations: asc, anterior sclerite; ey, eye; en, endopodite; hyp, hypostome; lm, lamellae; pb, podomere boundary; taf, transverse anterior fold. Figure 9. Incomplete specimens of Helmetia expansa. A, ROMIP 66305. B, ROMIP 66306. C, ROMIP 66318. D, ROMIP 66312. E, ROMIP 66315. F, ROMIP 61136. G, USNM 213815. H, USNM 189225. I, ROMIP 66317. Abbreviations: anl, anterolateral spine; ant, antenna; ey, eye; m, moult; mey, medial eyes; th, thorax; ts, terminal spine.
Figure 9. Incomplete specimens of Helmetia expansa. A, ROMIP 66305. B, ROMIP 66306. C, ROMIP 66318. D, ROMIP 66312. E, ROMIP 66315. F, ROMIP 61136. G, USNM 213815. H, USNM 189225. I, ROMIP 66317. Abbreviations: anl, anterolateral spine; ant, antenna; ey, eye; m, moult; mey, medial eyes; th, thorax; ts, terminal spine. Figure 10. Digestive system in Helmetia expansa ROMIP 66311. A, complete specimen in dorsal view. B, magnification of paired digestive structures from box 1 of A. C, magnification of digestive structure from box 2 of B. D, magnification of digestive structure from box 3 of B.
Figure 10. Digestive system in Helmetia expansa ROMIP 66311. A, complete specimen in dorsal view. B, magnification of paired digestive structures from box 1 of A. C, magnification of digestive structure from box 2 of B. D, magnification of digestive structure from box 3 of B. Figure 12. Arthroaspis bergstroemi from the Sirius Passet with gut preservation. A, C, MGUH 30397. B, D, MGUH 30382, holotype. Abbreviations: ex, exopodite; mg, midgut gland. Photo credit Martin Stein (Stein et al., 2013).
Figure 12. Arthroaspis bergstroemi from the Sirius Passet with gut preservation. A, C, MGUH 30397. B, D, MGUH 30382, holotype. Abbreviations: ex, exopodite; mg, midgut gland. Photo credit Martin Stein (Stein et al., 2013). Figure 11. Phylogeny of Artiopoda. A, maximum parsimony analysis with equal weights, strict consensus of 48 most parsimonious trees (tree length = 331 steps, CI = 0.41, RI = 0.71); Bremer support values in bold. B, maximum parsimony analysis with k = 5, strict consensus of 49 most parsimonious trees (CI = 0.41, RI = 0.71); node support values in bold as group present/contradicted frequency differences. C, consensus tree from Bayesian analyses in MrBayes using Mk model, four chains, 5,000,000 generations, 1/1000 sampling, 25% burn-in with 3610 retained samples. Posterior probability values in bold at nodes.
Figure 11. Phylogeny of Artiopoda. A, maximum parsimony analysis with equal weights, strict consensus of 48 most parsimonious trees (tree length = 331 steps, CI = 0.41, RI = 0.71); Bremer support values in bold. B, maximum parsimony analysis with k = 5, strict consensus of 49 most parsimonious trees (CI = 0.41, RI = 0.71); node support values in bold as group present/contradicted frequency differences. C, consensus tree from Bayesian analyses in MrBayes using Mk model, four chains, 5,000,000 generations, 1/1000 sampling, 25% burn-in with 3610 retained samples. Posterior probability values in bold at nodes. Figure 14. Reconstruction of the Burgess Shale concilitergan Helmetia expansa.Artwork by Marianne Collins.
Figure 14. Reconstruction of the Burgess Shale concilitergan Helmetia expansa.Artwork by Marianne Collins.
Losso, S. R., Caron, J. B., & Ortega-Hernández, J. (2025).
Helmetia expansa Walcott, 1918 revisited – new insights into the internal anatomy, moulting and phylogeny of Conciliterga. Journal of Systematic Palaeontology, 23(1). https://doi.org/10.1080/14772019.2025.2468195
Copyright: © 2027 The authors.
Published by Informa UK Limited. Open access.
Reprinted under a Creative Commons Attribution 4.0 International license (CC BY 4.0)













In short, there was nothing fundamentally different or exceptional about the biological processes operating before, during, or after the Cambrian Period.
This case also illustrates the inherently provisional nature of scientific understanding. Early interpretations of Helmetia expansa, based on limited and poorly preserved material, left its classification unresolved for over a century. The reassessment and reclassification, made possible by new techniques and better-preserved specimens, demonstrate how science refines and, when necessary, replaces earlier conclusions in light of improved evidence. Far from being a weakness, this self-correcting process is one of science’s greatest strengths—ensuring that our models of the natural world remain responsive to new discoveries and better data.
Advertisement
Amazon

Amazon

Amazon

Amazon

Amazon

Amazon

Amazon

Amazon

All titles available in paperback, hardcover, ebook for Kindle and audio format.
Prices correct at time of publication. for current prices.

No comments :
Post a Comment
Obscene, threatening or obnoxious messages, preaching, abuse and spam will be removed, as will anything by known Internet trolls and stalkers, by known sock-puppet accounts and anything not connected with the post,
A claim made without evidence can be dismissed without evidence. Remember: your opinion is not an established fact unless corroborated.