 Having blogged a couple of days ago about the role of mimicry in evolution I decided to look more closely at the subject, especially the widespread mimicry found in butterflies. The results are fascinating.
Having blogged a couple of days ago about the role of mimicry in evolution I decided to look more closely at the subject, especially the widespread mimicry found in butterflies. The results are fascinating.But before I get on to that I'll just deal quickly with another aspect to evolution - speciation - following on from something I mentioned in the same blog. I pointed out how mimicry involved a two species both of which are prey to the same predator and where at least one of them is toxic or harmful to the predator.
I mentioned that the selection pressure for one species to become more and more like the harmful one depends on the presence of the predator in the local environment but the species range may well include areas with different predators, different comimics or indeed the absence of one or both. In this case, and in that part of the range there may be no selection pressure and no particular advantage in adopting the colour pattern of the toxic or harmful species, and there may even be a disadvantage because a different predator may not have any aversion to the mimicked colour.
So, let's take a hypothetical example of a widespread species coexisting in part of its range (we'll call it Area A) with both a toxic comimic and a predator with an aversion to the comimic. In a different part of the range (which we'll call Area B) there may be a different predator and the comimic may be absent. Yet there is nothing to prevent the genes for mimicry flowing from Area A into Area B where they might well act as advertisements rather than deterrents. After all, what's the point of protective mimicry if it isn't conspicuous?
In Area B then, these genes could be being removed from the gene-pool by Natural Selection and there would be a distinct advantage in any mutations which prevented the population in Area B from acquiring genes for mimicry. For example, individuals which selected against potential mates with the mimicry pattern would tend to produce more descendants with that selection preference than those which didn't discriminate, so sex selection and mating displays would come to feature in Area B. These genes would in turn be detrimental as they flowed back to Area A because their carriers would now have genes preventing their offspring acquiring the protective mimicry in Area A.
The environment, together with Natural Selection, would have created the conditions in which it is to both colour varieties' advantage to prevent interbreeding and to evolve barriers to it. In this situation we would expect the two gene-pools to quickly become effectively isolated from one another, or, to put it in taxonomic terms, two new species would have diverged from one original species.
This of course is a simplistic scenario. In reality, and especially with a widespread population, the situation may be far more complex with more than one comimic presenting different levels of risk to predators, more than one predator, overlapping predator ranges, etc, etc., so the result might be a whole range of intermediates, distinct colour forms predominating in different parts of the range and yet not enough environmental pressure for barriers to gene flow to arise.
Creationists tell us that it is impossible for a new species to arise by evolution because they have been fed the lie by creationist pseud-scientists that there is a fundamental difference in the mechanism by which a species changes over time due to changes in its genome, and the mechanism by which gene-pools become effectively isolated with barriers to interbreeding. I would like a creationists therefore to explain to me how the mechanism I have described above by which genes for mimicry are acquired in one part of the range and barriers to the spread of those genes into a different part of the range is fundamentally different and why one is perfectly possible and the other is impossible. During what stage does the mechanism change as the species evolves from allowing full interbreeding to full gene-pool isolation and how does that mechanism then become impossible retrospectively when previously it was perfectly possible? Surely there is a creationist site where this is fully explained, isn't there?
Anyway, on the subject of mimicry in butterflies, scientists have recently discovered that, at least in the Asian swallowtail butterfly, Papilio polytes, wing shape, colouration and pattern are all controlled by a single 'supergene', i.e., a group of genes all physically grouped together on a chromosome making cross linking less likely and so tending to all be inherited together as a single package. Moreover, they have discovered that this supergene is a modified form of another supergene called the doublesex gene which controls sexual dimorphism in butterflies. At some point in their evolutionary history, probably close to the point of divergence of butterflies from the silk moths this gene became doubled so one copy was free to mutate and evolve without being detrimental to the doublesex gene's function.
Abstract
One of the most striking examples of sexual dimorphism is sex-limited mimicry in butterflies, a phenomenon in which one sex—usually the female—mimics a toxic model species, whereas the other sex displays a different wing pattern. Sex-limited mimicry is phylogenetically widespread in the swallowtail butterfly genus Papilio, in which it is often associated with female mimetic polymorphism. In multiple polymorphic species, the entire wing pattern phenotype is controlled by a single Mendelian 'supergene'. Although theoretical work has explored the evolutionary dynamics of supergene mimicry there are almost no empirical data that address the critical issue of what a mimicry supergene actually is at a functional level. Using an integrative approach combining genetic and association mapping, transcriptome and genome sequencing, and gene expression analyses, we show that a single gene, doublesex, controls supergene mimicry in Papilio polytes. This is in contrast to the long-held view that supergenes are likely to be controlled by a tightly linked cluster of loci. Analysis of gene expression and DNA sequence variation indicates that isoform expression differences contribute to the functional differences between dsx mimicry alleles, and protein sequence evolution may also have a role. Our results combine elements from different hypotheses for the identity of supergenes, showing that a single gene can switch the entire wing pattern among mimicry phenotypes but may require multiple, tightly linked mutations to do so.
So, at least in this swallowtail butterfly, a single supergene controls the form of mimicry the carrier will have and, because this is tied in with the gene controlling gender expression, mimicry is limited to females.
| Papilio Polytes Morph | Comimic Species | ||||
|---|---|---|---|---|---|
| No comimic species. Sexes are alike. | ||||
|
| ||||
|
|





Distribution of the different morphs of Papilio Polytes females corresponds closely with that of the comimic species of Pachliopta.
|
|

In this group from South America, each species of Heliconius has a corresponding comimic in the Mechanitis genus.
|
|

Note that each of the top three morphs of Pseudacraea eurytus has a mimic, even mimicking the sexual dimorphism seen in the top two. This linkage of sexual dimorphism and mimicry type reflects the fact that the supergenes for both are closely related and controlled in the same way. A female will also inherit the female mimicry supergene and the male will inherit the male one. These supergenes, although in different species have remained relatively stable in the way they are arranged and in how they express since early in the evolutionary history of butterflies. This can also mean that there is not a whole array of genes which need to evolve as mimicry develops but the entire change can be due to a small number of changes or even a single 'switch'.
Mimicry is by no means confined for insects or course. The following shows Müllerian mimicry in poisonous South American frogs. The top row are all members of the same species (Dendrobates imitator) which have evolved in different areas to resemble other poisonous species.
 |  |  |
| Above: regional morphs of Pseudacraea eurytus | ||
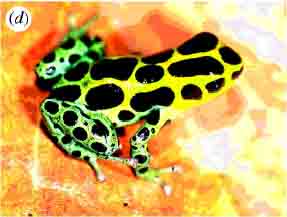 |  |  |
| Dendrobates variabilis | Dendrobates fantasticus | Dendrobates ventrimaculatus |
With Mülarian mimicry, two harmful species come to resemble one another as a kind of spread bet. The display is conspicuous and each benefits from a predator's aversion to the other.
 |
| Coralsnake and a scarlet kingsnake. Source: http://www.knewance.com/comparisons/coral-snake-v-milk-snake-v-scarlet-kingsnake.html |
There is of course absolutely no way evolved mimicry can be fitted into an intelligent design model because it is entirely dependent on wasteful competition, predation and avoidance. There is no intelligent reason to design a predator to prey on one species (even if we forget the supposed benevolence) and then design the prey to deter the predator from eating it, and without this need for deterrence there is no reason for the prey to be toxic in the first place so no benefit would accrue from mimicking it.
The only rational explanation for mimicry in living things is as the result of a mindless, purposeless, evolution over time directed only by an unemotional natural selection which results in the genes of those which leave the most descendants coming to predominate in the species local gene-pool. In their advocacy of intelligent design, creationists pseudo-scientists betray one of two things: an ignorance of the subject on which they expound with feigned expertise or a disregard for truth, honesty and integrity.
As even devout Old Earth Creationist, Francis Collins pointed out in an otherwise thoroughly dishonest book, The Language of God, Young Earth Creationism (i.e, those who advocate ID and try to have it inserted into school science curricula in pursuit of an extremist political agenda) have reached a point of bankruptcy, both in its science and in its theology.

I read the other day your blog article Copycat Evolution, Rosa. See: http://rosarubicondior.blogspot.se/2014/07/copycat-evolution.html
ReplyDeleteIn that article you told me and your other followers about two main types of mimicry. Let me quote from your article:
There are in fact two main types of mimicry recognised by biologists, although the distinction is quite technical and is blurred anyway. For all practical purposes, they can both be regarded as forms of the same thing. They are known to biology as:
[1] Batesian mimicry. Named after the English naturalist Henry Walter Bates. This is where a harmless species evolves to resemble a harmful species if it and the harmful species share a common predator.
[2] Müllerian mimicry. Named after the German biologist Fritz Müller. This is where two harmful species evolve to resemble one another if they both share a common predator.
Therefore I suspect a "lapsus manus" or "lapsus calami" in today's new blog article, since you here call Fritz Müller's type of mimicry a "Mülarian" mimicry instead of a "Müllerian one".
Now this is of course but a trifle, but I mention it because you are known for your acribia or scrupulousness.
BTW: As is my wont I'd like to recommend a site where some basic concepts in evolution are explained: http://darwin200.christs.cam.ac.uk/pages/index.php?page_id=g6 .
Just this URL leads you to an explanation of the mimicry concept in a fairly funny way. I quote some sentences from that article:
Better 5% Turd Than No Turd At All. (...) The key is that evolution takes small steps over time. The ancestral Eastern tiger swallowtail caterpillar probably looked nothing like a bird turd, and its predators probably completely ignored bird turd as a source of food. In this population any mutation that occurred which made one caterpillar look 2% more turd-like than the others would give that caterpillar a greater chance of survival, as predators would be a little more likely to discount it as a food source. That caterpillar would grow up, reproduce, its offspring would look 2% like a turd and the cycle repeats. Eventually the 2% turd mutation would spread throughout the population. As it becomes more common there would be selective pressure on the predator to identify things that look slightly poo-like as potential food sources. This means that if a second mutation occurred which made the caterpillar look 5% like a turd it would then spread in the same way. Selection would then favour birds able to identify these caterpillars. This could then repeat until we end up with caterpillars that look almost 100% like a bird poo and predators that can tell the difference between a 99% bird poo-like caterpillar and a real bird poo.
Another evolutionary concept is discussed and explained here: http://darwin200.christs.cam.ac.uk/pages/index.php?page_id=g2 . That article is called "Irreducibly Complex?".
I strongly recommend all creationists to read not only Rosa Rubicondior's articles about evolution here on her fine and knowledgeable blog but also the articles on this other site.
Thanks. I'll correct the typo. :-)
Delete