In an unguarded moment, for which he has had to apologize, Pope 'Mr. Nice Guy' Francis, revealed the hypocritical difference between his private and public declarations.
Trying to give the Catholic Church a more tolerant and understanding face, in place of the bigoted and judgmental church of his predecessors, that had been driving decent people away, and aware that many Catholic Priests are practicing homosexuals, Pope Francus has previously advocated tolerance and understanding, whilst falling short of allowing same-sex marriages in Catholic churches and ordaining openly gay priests.
But that mask of kindly affability and tolerance was revealed to be a lie intended to deceive when Francis let slip his true feelings when asked if he would allow gay men to enter seminary to be trained and eventually ordained as Catholic priests.
He replied (in Italian):
Seminaries are already too full of “frociaggine”.
This translates as 'faggotiness' and is an especially abusive term for gay people in Italy.
According to recent findings by researchers at the Max Delbrück Center and the European Molecular Biology Laboratory, just published, open access, in the journalNature Genetics,
it all depends on which of your cells we look in.
You might think that the replication process, as our cells divide during growth and development, and to replace old, damaged and worn-out cells, would be perfect if designed by a perfect, omniscient designer, but apparently not. And as we age, the mistakes accumulate until we are a mosaic of different genomes, contributing to diseases caused by genetic abnormalities, including cancers.
On the other hand, this is exactly the sort of thing that a utilitarian, evolutionary process, where a suboptimal process will be retained if it gives more descendants that what went before, can be predicted to produce.
How the team discovered this is described in a news release from the Max Delbrück Center:
Fig 4. Taphonomic modification on gomphothere bones.
A-B. Astragalus (TT3-E5-N18-06) with tooth marks in ventral and dorsal view (discontinued line indicates the original extension of the specimen); C. caudal vertebra (TT3-E4-N18/19-02) with a single cutmark; D. caudal vertebra (TT3-E5/F5-N18-53) with several parallel cutmarks.
Between 2,000 and 2,500 years before creationists believe their little god created a small universe consisting of a single, small flat planet with a dome over it, in the Middle East, human beings were hunting a now-extinct elephant on the shores of Tagua Tagua Lake in Chile, South America. By then, humans had migrated out of Africa, spread across Asia to the land bridge between Siberia and Alaska, and from there into North America, down through the Panamanian Isthmus, or by coastal spread, down to South America. Elephants had preceded them probably by several thousand years.
The evidence for this is presented in a paper in PLOS ONE by Rafael Labarca of the Pontifical Catholic University of Chile and colleagues.
According to information made available by PLOS ahead of publication, and reported in Science Daily:
The origin of the so-called 'German' cockroach, which has nothing especially to do with Germany, has been something of a mystery, as it is unknown outside human habitation. It is what is known as an obligate, commensal species which, like the barn swallows, house mice, human lice and bed bugs, has co-evolved with human culture to live in and on humans and their buildings.
They neatly illustrate how speciation occurs when a population learns to exploit a new resource and then becomes genetically isolated from its parent population and so evolves over time into a new species.
Creationists will tell you that the Bible is the 'complete and inerrant' record of the history of life on planet earth and an accurate scientific description of the life on it, and yet it's so inadequate that they've had to supplement it with all manner of fanciful additions to make any sense of it and to make it look slightly more connected to reality.
They then get themselves into a hopeless muddle trying to define a 'kind’, so it conforms to their nonsensical taxonomy. They will tell you one 'kind' can't interbreed with another 'kind' and then need to use a different definition of 'kind' when you mention hybrids, ring species and other evidence of slow divergence over time as barrier to hybridization and genetic remixing evolve, before what science recognises as genetically distinct genera and species.
In any social media group or creationists disinformation site, you'll see a typical double-think approach to evolution: on the one hand we'll be assured that there is no evidence for evolution (based on some childish parody definition of evolution); on the other hand, you'll be assured that, after two of each 'kind' of animal got off the mythical Ark, they underwent a period of warp-speed evolution, producing several, entirely new taxons each generation in different parts of their range to give us the millions of different families, species, sub-species and varieties we see today, which, coincidentally, no-one reported noticing, not even the people who left rock and cave paintings recording what they saw in the world around them several thousand years ago.
But of course, evolution is what it has always been since biologists gained insight into how life changes in response to environmental change - change in the frequency of different alleles in the population over time.
These changes may or may not include new structures or morphology that taxonomists base their classifications on, and they may or may not include changes in coloration (which is a morphological change just as much so as is a new structure).
Here, for example, is the recent discovery by a team lead by Zachariah Gompert of the Department of Biology, Utah State University, Logan, UT, USA, that the stick insect species, Timema cristinae, has had several predictable, repeats of the same pattern of evolution in response to cyclic changes in their environment.
The species occurs in two colour forms - a uniform green, which cryptically camouflages it as leaves of the Californian lilac shrub, Ceanothus spinosus, and a green-striped form which cryptically camouflages it as the leaves of the chamise shrub, Adenostoma fasciculatum.
What the team discovered is the subject of both an open-access paper in Science and a news release from Utah State University:
A longstanding debate among evolutionary scientists goes something like this: Does evolution happen in a predictable pattern or does it depend on chance events and contingency? That is, if you could turn back the clock, as celebrated scientist Stephen Jay Gould (1941-2002) described in his famous metaphor, “Replaying the Tape of Life,” would life on Earth evolve, once again, as something similar to what we know now, or would it look very, very different?
“If you frame it as an either/or question, it’s too simplistic,” says Utah State University evolutionary biologist Zachariah Gompert. “The answer isn’t ‘completely random’ or ‘completely deterministic and predictable.’ And yet, examining short time scales, we can find predictable, repeatable evolutionary patterns.”
Gompert and colleagues report evidence of repeatable evolution in populations of stick insects in the May 24 online edition of the American Association for the Advancement of Science’s journal Science Advances. Contributing authors on the paper include Gompert’s long-time collaborator Patrik Nosil and other researchers from France’s University of Montpelier, Brazil’s Federal University of São Paulo, the University of Nevada, Reno and Notre Dame University. The research is supported by the National Science Foundation and the European Research Council.
The team examined three decades of data on the frequency of cryptic color-pattern morphs in the stick insect species Timema cristinae in 10 naturally replicate populations in California. T. cristinae is polymorphic in regard to its body color and pattern. Some insects are green, which allows the wingless, plant-feeding insect to blend in with California lilac (Ceanothus spinosus) shrubs. In contrast, green striped morphs disappear against chamise (Adenostoma fasciculatum) shrubs.
Hiding amongst the plants is one of T. christinae’s key defenses as hungry birds, such as scrub-jays, are insatiable predators of the stick insects.|
“Bird predation is a constant driver shaping the insects’ organismal traits, including coloration and striped vs. non-striped,” says Gompert, associate professor in USU’s Department of Biology and the USU Ecology Center. “We observed predictable ‘up-and-down’ fluctuations in stripe frequency in all populations, representing repeatable evolutionary dynamics based on standing genetic variation.”
He says a field experiment demonstrates these fluctuations involved negative frequency-dependent natural selection (NFDS), where cryptic color patterns are more beneficial when rare rather than common. This is likely because birds develop a “search image” for very abundant prey.
“At short time scales, evolution involving existing variations can be quite predictable,” says Gompert, who received a National Science Foundation CAREER grant in 2019 to support his research. “You can count on certain drivers always being there, such as birds feeding on the insects.”
But at longer time scales, evolutionary dynamics become less predictable.
“The populations might experience a chance event, such as a severe drought or a flooding event, that disrupts the status quo and thus the predictable outcomes,” Gompert says.
On long time scales, a new mutation in the species could introduce a rare trait, he says. “That’s about as close to truly random as you can get.”
“Rare things are easily lost by chance, so there’s a strong probability a new mutation could disappear before it gains a stronghold,” he says. “Indeed, another species of Timema stick insect that also feeds on chamise either never had or quickly lost the mutations making the cryptic stripe trait. Thus, the evolution of stripe is not a repeatable outcome of evolution at this long scale.”
Gompert notes replicated, long-term studies from natural populations, including research on the famous Darwin’s finches, are rare.
“Because most of this work is restricted to one or few populations, it is difficult to draw inferences on repeatability among multiple evolutionary independent populations,” he says. “Such studies are challenging to implement not only because they take concerted effort, but also because you can’t rush time.”
Abstract
The extent to which evolution is repeatable remains debated. Here, we study changes over time in the frequency of cryptic color-pattern morphs in 10 replicate long-term field studies of a stick insect, each spanning at least a decade (across 30 years of total data). We find predictable “up-and-down” fluctuations in stripe frequency in all populations, representing repeatable evolutionary dynamics based on standing genetic variation. A field experiment demonstrates that these fluctuations involve negative frequency-dependent natural selection (NFDS). These fluctuations rely on demographic and selective variability that pushes populations away from equilibrium, such that they can reliably move back toward it via NFDS. Last, we show that the origin of new cryptic forms is associated with multiple structural genomic variants such that which mutations arise affects evolution at larger temporal scales. Thus, evolution from existing variation is predictable and repeatable, but mutation adds complexity even for traits evolving deterministically under natural selection.
INTRODUCTION
The extent to which evolution is repeatable and predictable is central to understanding the role of determinism and chance in the history of life, with implications for both basic and applied science (1–4). These ideas are captured in Gould’s famous metaphor of “replaying the tape of life” (5). Gould argued that historical contingency and chance idiosyncrasies would result in different (i.e., non-repeatable) evolutionary outcomes if the history of life was to be replayed over and over again. However, others such as Morris (6) have argued that evolution is inherently predictable, and many examples of deterministic natural selection promoting repeatable and predictable elements to evolution do exist (3, 7). Thus, beyond spurring decades of debate, Gould’s metaphor has been usefully transformed into an empirical research program (1, 8).
Comparative phylogenetic studies of parallel evolution often support repeated outcomes driven by natural selection (1, 7, 9). However, inferring evolutionary processes and their interplay from such retrospective work can be challenging. For example, selection that fluctuates rapidly between time points can be misconstrued as evolutionary stasis if only end outcomes, or a few distant time points, are analyzed. Detailed studies of the fossil record and ancient DNA can help address these issues (10, 11), as can real-time studies of evolutionary dynamics. The latter are exemplified by studies of experimental evolution in replicated laboratory populations of microbes (12–14) and other organisms, such as insects (15, 16). This impressive body of work has revealed not only repeatable patterns of evolution by natural selection but also a role for the contingency of mutation.
In contrast to laboratory experimental evolution studies, replicated, long-term studies from natural populations are rare. Certainly, highly influential long-term studies of the predictability of evolution in the wild do exist, in finches and other birds (17–20), moths and butterflies (21–23), stickleback and guppy fish (24–27), sheep (28), fruit flies (29), and deer (30) (to name a few). For example, rare climatic events affect evolution in the famous Darwin’s finches (19, 20). However, most such work is restricted to one or few populations, making it difficult to draw inferences on repeatability among multiple evolutionarily independent populations (note the distinction between predictability within a population and repeatability among populations, Fig. 1). Thus, replicated studies of evolution in the wild are required to test the generality of the findings from microbes (12–14) and to help bridge now disparate laboratory and field studies. Such studies are challenging to implement not only because they take concerted effort but also because time cannot simply be sped up with more effort.
Fig. 1. Evolution in replicate long-term field populations of T. cristinae stick insects.
(A) Illustrations of the color-pattern morphs of T. cristinae. (B) Predictions of highly repeatable evolutionary dynamics over time. Each line represents a different population, each exhibiting predictable “up-then-down” fluctuations in trait or gene frequency over time. (C) Predictions of less repeatable evolutionary dynamics over time. In contrast to the panel to the left, each population exhibits different patterns of trait or gene frequency change over time. Note the distinction between predictability within any single time series (i.e., population or replicate) and repeatability among them. (D) Empirical variation in morph frequencies in T. cristinae between 1990 and 2023. The orange line (median) and shading [95% equal-tailed probability interval (ETPI)] represent yearly averages on the host plant Adenostoma. The blue line (median) and shading (95% ETPI) represent yearly averages on the host plant Ceanothus. (E) Population-specific morph frequency variation over time, representing replicate evolutionary dynamics (mean number of years per population = 14). Results are shown for the 10 core populations, i.e., replicates, that this study focuses on.
Here, we provide such a study on the basis of compiled data from 30 years of tracking morph frequencies across 10 replicate populations of a stick insect in the wild (Fig. 1). We integrate these data with field experiments, modeling, and genomic data to elucidate the processes driving evolutionary dynamics across timescales. In particular, our genomic analyses allow consideration of the contingency of mutation, which can add complexity and nuance relative to the sole consideration of evolution from standing genetic variation (31). Specifically, mutational dynamics can be related to the genetic architecture of traits, with consequences for the repeatability of evolution at the genetic versus phenotypic level (32, 33). Some predictions are as follows and illustrated in Fig. 2.
Fig. 2. Conceptual diagram illustrating the relationship between the genetic architecture of a trait and the repeatability of evolution at the genetic and phenotypic levels (where the tips of each phylogeny represent a different taxon/species).
(A) Hypothetical scenario where a trait (i.e., stripe) depends on a specific mutation. In this case, the evolution of stripe is contingent on the specific mutation occurring and thus occurs only once in the hypothetical evolutionary scenario depicted on the phylogeny. Thus, phenotypic evolution is not repeatable (it would, however, have a repeatable genetic basis if the trait evolved in multiple species). (B) An alternative scenario where stripe depends on a few loci. Here, the evolution of stripe is still contingent on a small number of specific mutations occurring and this might make the trait evolve less repeatedly. Moreover, in such cases, evolution might (or might not) be dependent on the order in which the mutations occur (i.e., some mutations might only be beneficial if another mutation has already occurred). (C) A third alternative scenario where stripe is affected by many mutations (i.e., is polygenic). In this case, stripe does not depend on a specific, unlikely mutation, and is unlikely to depend on the order of mutations. This genetic architecture thus makes the repeated evolution of stripe much more likely, occurring in all four species in the hypothetical example. However, the genetic basis of stripe varies among the species, and, thus, the genetic basis of stripe is not repeatable (different combinations of alleles, i.e., mutations, can generate a stripe). In all diagrams, + and − symbols denote alternative alleles, with + contributing to the stripe phenotype.
When traits are controlled by simple genetic architectures composed of one or few loci of large effect, then individual mutations can strongly affect evolutionary dynamics (4, 34). In such cases, individual mutations “matter” and mutation can be an important consideration for understanding evolution. For example, such architectures offer few mutational targets, making mutations that improve fitness rare (4, 34). Moreover, if there are strong interactions between loci (i.e., epistasis), then only a few genetic combinations may work well together to increase fitness. In other words, there are few mutational sequences or “paths” that adaptive evolution can actually take (4, 34–36). Thus, traits controlled by few loci may be challenging to evolve de novo and thus show modest repeatability in their origin over time. However, when such traits do evolve, they will have a predictable, repeated genetic basis (i.e., using the same, few loci and mutations that increase fitness). These predictions could be different for more polygenic architectures composed of many loci with smaller effects (37). Here, no individual mutation has a strong effect on a trait, many loci have redundant effects, and path dependency is unlikely to force a particular mutational sequence to be used to evolve higher fitness. Thus, traits controlled by many loci may evolve repeatedly because they can use a wide range of mutational variation. However, when they do so, a different set of loci and mutations is likely to be used in each instance, making evolution at the genetic level not very repeatable (37).
We acknowledge that there can be much nuance beyond the stylized ideas and predictions noted above [e.g., see (38)]. However, these ideas, nonetheless, illustrate why it can be informative to study the genetic architecture of traits that are being analyzed for their repeatability, particularly to understand the dynamics of evolution from standing variation versus new mutation. In this context, we here combine our time-series and experimental work with genomic analyses of the genetic architecture of cryptic color pattern. Our results reveal clear similarities but also differences with past work focused on body color (rather than pattern, see below for details). Our integrative approach leads to a more complete understanding of the repeatability of evolution than would be possible by studying selection or mutation in isolation.
Ecology and genetics of Timema stick insects
Our study system is Timema stick insects (Fig. 1), a genus of wingless, plant-feeding insects found throughout southwestern North America (39). We focus primarily (but not exclusively) on Timema cristinae, which exhibits three highly heritable morphs and uses two primary host plant species (40, 41). Two of these morphs have diverged in frequency between host species due to strong divergent selection imposed by visual predators such as lizards and birds (42, 43). Specifically, a green unstriped color-pattern morph (green morph hereafter) is cryptic on the broad leaves of Ceanothus. In contrast, the striped color-pattern morph (a green morph that also bears a white, longitudinal stripe on its dorsal surface) is cryptic on the thin needle-like leaves of Adenostoma. Thus, each morph is generally more common on the host on which it is more cryptic, which led to a body of past work on the potential for divergent adaptation between hosts to drive ecological speciation (41, 44).
However, we stress that polymorphism is maintained such that both green and striped morphs occur in most populations. This variation is maintained due to negative frequency-dependent selection (NFDS) and gene flow, creating a mosaic of variation in morph frequencies across the landscape, both within and among populations (45, 46). Specifically, the frequency of the striped morph varies within local populations ranging from low (near zero), to intermediate, to high (near one) but is rarely truly fixed (Fig. 1). This provides the requisite variation to study changes in the frequency of green versus striped morphs over time, which we focus on here. There is also a third darkly colored or melanic morph that is rarer and found at comparable frequencies between the two hosts. The melanic morph is not specifically adapted to either host (40, 47) does not fluctuate strongly in frequency over time and does not appear subject to NFDS (40, 45, 47). Thus, we focus first on testing for repeatable fluctuations in color-pattern morphs but return to melanism later in this study when considering longer timescales. We also return to the relationship between polymorphism and speciation at the end of our study.
Past observations and a manipulative field experiment documented changes in color-pattern morph frequencies that were highly predictable due to NFDS (i.e., a fitness advantage to rare forms) (45). This likely occurs due to birds switching search images to hunt for common prey items (48–51). Thus, increases in the frequency of the striped morph one year were reliably followed by decreases the following year and vice versa. This past work focused on changes in a single 18-year time series (i.e., a single population or “play of the tape”) because substantial long-term data from other populations did not exist at the time. Thus, past work could not address the issue of repeatability or “replays,” i.e., among replicate populations, per se. We thus focus here on the repeatability of evolution among populations.
We report results from 30 years of data collection on morph frequencies in T. cristinae, representing 692 year-by-host-by-locality estimates of morph frequency derived from 48,349 individuals (Fig. 1 and data S1). Most critically, these data represent 10 localities with at least a decade of the data required to test for NFDS (see table S1; mean, 14 years; maximum, 22 years). These 10 localities were chosen to represent replicates, for example, because of little to no gene flow among them such that each locality undergoes independent yearly changes in morph frequency. Supporting this claim, we emphasize that our 10 localities are geographically separated (fig. S1), generally by several kilometers, yet the average per-generation dispersal distance based on a mark-recapture study is only 12 m (52). This makes it very unlikely that there is sufficient migration between our localities to generate detectable morph frequency fluctuations. Molecular data further support limited gene flow between our localities and thus their evolutionary independence (especially in the context of yearly changes in morph frequency). For example, genetic data have shown that even parapatric populations that are directly adjacent to one another exchange only a few migrants per generation, and all of our study localities here are geographically separated from each other such that gene flow among them is even lower (fig. S1) (53, 54). Thus, each locality acts as a replicate or replay for analyzing the repeatability of evolution, particularly because the distance (i.e., kilometers) typically separating each locality make it likely that different bird and lizard individuals hunt insects at each locality.
Notably, T. cristinae is univoltine with nonoverlapping generations. Thus, each year of data represents a generation, with evolution occurring between each pair of years (the insects diapause as eggs through autumn and winter, hatch in spring, and mate and die in early summer, repeating this cycle each generation). Despite these data representing an appreciable effort, the timescales involved are very different from those associated with Gould’s metaphor. We thus also consider mutations affecting color pattern over deeper evolutionary time (55).
Here, the genetic architecture of color pattern is relevant, for reasons outlined above and in Fig. 2. Specifically, we here evaluate the extent to which stripe depends on specific mutations that could make the repeated evolution of the trait less likely. Past work suggests that pattern is controlled by one or few loci on chromosome 8 (chr8 hereafter) (40, 47, 56). However, the number, effect sizes, and physical distributions of the loci involved remains unclear. Specifically, past work described a region of chr8 named the Mel-Stripe locus that harbors complex structural variation that explains most of the variation in body color (a large inversion and deletion distinguish green versus melanic morphs, indicative of suppressed recombination and opening the potential for linked selection) and is also partially associated with color pattern (striped versus green). However, whether one or multiple regions of Mel-Stripe or even other loci are associated with pattern remains unclear due to (i) the focus of past work on color (not pattern) and (ii) the use of a fragmented reference genome. Here, we integrate better chromosome-level genome assemblies with genome-wide association (GWA) mapping to show that color pattern is actually associated with multiple different structural variants within the Mel-Stripe locus, as well as a chromosomal inversion in a region not associated with color or pattern in past work (i.e., the “Pattern” locus, which is reported here for the first time). We conclude our study by discussing what these genetic details tell us about the repeatability of evolution.
Fig. 9. Two solutions to the problem of crypsis on Adenostoma (in T. cristinae versus T. podura) and an experimental test of which solution may offer higher fitness.
(A) Conceptual overview of known patterns of crypsis. Both T. cristinae and T. podura use Ceanothus and Adenostoma as host plants and both exhibit a green unstriped morph that is cryptic on Ceanothus. In contrast, the two Timema species have evolved different morphs that are cryptic on Adenostoma, the striped morph in T. cristinae (left) and a melanistic gray/brown morph in T. podura (right). (B) Block-specific results of the mark-recapture field experiment testing the fitness of striped versus melanistic morphs of T. cristinae on Adenostoma. Shown is the number of each morph recaptured in each block (i.e., replicate), demonstrating higher recapture and thus fitness of the striped morph. (C) Average fitness of each morph across the entire experiment with the posterior distributions summarized in box plots (boxes denote the median, first and third quartiles of the posterior with whiskers extending to the minimum and maximum or 1.5 times the interquartile range) and individual parameter values sampled from the posterior overlain as points.
In summary, the population of the different morphs changes according to the strength of the selectors in the environment, in a predictable way, at least in the short-term, predictable being in the way the Theory of Evolution by Natural selection predicts. There is no suggestion here that some other, supernatural, force is involved in the process or that the biologists think the TOE is inadequate for explaining the observations. Quite the contrary, in fact, the TOE is confirmed by the observations.
For the first time in the history of astronomy, researchers at the Niels Bohr Institute have witnessed the birth of three of the universe's absolute earliest galaxies, somewhere between 13.3 and 13.4 billion years ago.
Imagine you could look back in time to witness events soon after the Big Bang. How devastating would that be to Ken Ham's strategy of teaching unfortunate children not to enquire how scientists know stuff but to dismiss what they say with the smugly condescending sneer, "Were you there?" as though the only way to know anything is to witness it yourself.
But with the ability to look back in time and actually witness events, and to confirm that those events are exactly as the science predicts based on the normal logical deductions that normal people use - like knowing that your great grandparents had sex, even though no-one witnessed them and they left no record, other than a children born 40 weeks later. According to Ham, there is no way to know that your great grandparents had sex because you weren't there to witness it, so the claim they did has no scientific basis [sic].
So, a child born 40 weeks later could just as well be the result of magic as the result of your great grandparents having sex.
Yep! That's how silly Ham's strategy is and how stupid he has made the willing dupes who use it, sound.
But, to get back to the point of actually looking back in time to witness events 13.4 billion years ago. How is that even possible?
It's possible because of the finite speed of light. When we look deep into space, we aren't seeing events and objects as they are today but as they were when the light reaching our eyes or the detectors on telescopes started out. The further we look into space, the further back in time we are looking, so, when we see objects and events that are 13.4 billion lightyears away, we are seeing them as they were 13.4 billion years ago
And what we see are the first galaxies forming.
So yes, we were there, just as we were there when something happened in the same room. The only difference being the time it took the light to reach our eyes - a few picoseconds or a few billion years. We no less witness an actual event because it took billions of years for information about it to reach us, as we witness an event a few picoseconds ago.
How the scientists at the Cosmic Dawn Center at the University of Copenhagen’s Niels Bohr Institute is explained in a University of Copenhagen Faculty of Science news release, and in a paper published today in Science:
THE UNIVERSE Using the James Webb Space Telescope, University of Copenhagen researchers have become the first to see the formation of three of the earliest galaxies in the universe, more than 13 billion years ago. The sensational discovery contributes important knowledge about the universe and is now published in the prestigious journal Science.
For the first time in the history of astronomy, researchers at the Niels Bohr Institute have witnessed the birth of three of the universe's absolute earliest galaxies, somewhere between 13.3 and 13.4 billion years ago.
The discovery was made using the James Webb Space Telescope, which brought these first 'live observations' of formative galaxies down to us here on Earth.
Through the telescope, researchers were able to see signals from large amounts of gas that accumulate and accrete onto a mini-galaxy in the process of being built. While this is how galaxies are formed according to theories and computer simulations, it had never actually been witnessed.
You could say that these are the first 'direct' images of galaxy formation that we’ve ever seen. Whereas the James Webb has previously shown us early galaxies at later stages of evolution, here we witness their very birth, and thus, the construction of the first star systems in the universe.
Assistant Professor Kasper Elm Heintz, lead author
Cosmic Dawn Center, Copenhagen, Denmark.
Niels Bohr Institute,
University of Copenhagen, Copenhagen, Denmark.
Today, the study has been published in the esteemed scientific journal Science.
HOW THEY DID IT
Researchers were able to measure the formation of the universe’s first galaxies by using sophisticated models of how light from these galaxies was absorbed by the neutral gas located in and around them. This transition is known as the Lyman-alpha transition.
By measuring the light, the researchers were able to distinguish gas from the newly formed galaxies from other gas. These measurements were only possible thanks to the James Webb Space Telescope’s incredibly sensitive infrared spectrograph capabilities.
Galaxies born shortly after the Big Bang
The researchers estimate the birth of the three galaxies to have occurred roughly 400-600 million years after the Big Bang, the explosion that began it all. While that sounds like a long time, it corresponds to galaxies forming during the first three to four percent of the universe's 13.8-billion-year overall lifetime.
Shortly after the Big Bang, the universe was an enormous opaque gas of hydrogen atoms – unlike today, where the night sky is speckled with a blanket of well-defined stars.
During the few hundred million years after the Big Bang, the first stars formed, before stars and gas began to coalesce into galaxies. This is the process that we see the beginning of in our observations.
Associate Professor Darach Watson., co-author.
Cosmic Dawn Center, Copenhagen, Denmark.
Niels Bohr Institute,
University of Copenhagen, Copenhagen, Denmark.
The birth of galaxies took place at a time in the history of the universe known as the Epoch of Reionization, when the energy and light of some of the first galaxies broke through the mists of hydrogen gas.
It is precisely these large amounts of hydrogen gas that the researchers captured using the James Webb Space Telescope’s infrared vision. This is the most distant measurement of the cold, neutral hydrogen gas, which is the building block of the stars and galaxies, discovered by scientific researchers to date.
ABOUT THE EARLY UNIVERSE
The universe began its “life” about 13.8 billion years ago in an enormous explosion – the Big Bang. The event gave rise to an abundance of subatomic particles such as quarks and electrons. These particles aggregated to form protons and neutrons, which later coalesced into atomic nuclei. Roughly 380,000 years after the Big Bang, electrons began to orbit atomic nuclei, and the simplest atoms of the universe gradually formed.
The first stars were formed after a few hundred million years. And within the hearts of these stars, the larger and more complex atoms that we have around us were formed.
Later, stars coalesced into galaxies. The oldest galaxies known to us were formed about 3-400 million years after the Big Bang. Our own solar system came into being about 4.6 billion years ago – more than 9 billion years after the Big Bang.
Adds to the understanding of our origins
The study was conducted by Kasper Elm Heintz, in close collaboration with, among others, research colleagues Darach Watson, Gabriel Brammer and PhD student Simone Vejlgaard from the Cosmic Dawn Center at the University of Copenhagen’s Niels Bohr Institute – a center whose stated goal is to investigate and understand the dawn of the universe. This latest result brings them much closer to doing just that.
The research team has already applied for more observation time with the James Webb Space Telescope, with hopes of expanding upon their new result and learning more about the earliest epoch in the formation of galaxies.
For now, this is about mapping our new observations of galaxies being formed in even greater detail than before. At the same time, we are constantly trying to push the limit of how far out into the universe we can see. So, perhaps we’ll reach even further.
says Simone Vejlgaard, co-author.
Cosmic Dawn Center, Copenhagen, Denmark.
Niels Bohr Institute,
University of Copenhagen, Copenhagen, Denmark.
According to the researcher, the new knowledge contributes to answering one of humanity’s most basic questions.
One of the most fundamental questions that we humans have always asked is: 'Where do we come from?'. Here, we piece together a bit more of the answer by shedding light on the moment that some of the universe’s first structures were created. It is a process that we’ll investigate further, until hopefully, we are able to fit even more pieces of the puzzle together
Associate Professor Gabriel Brammer, co-author
Cosmic Dawn Center, Copenhagen, Denmark.
Niels Bohr Institute,
University of Copenhagen, Copenhagen, Denmark.
The study was conducted by researchers Kasper E. Heintz, Darach Watson, Gabriel Brammer, Simone Vejlgaard, Anne Hutter, Victoria B. Strait, Jorryt Matthee, Pascal A. Oesch, Pall Jakobsson, Nial R. Tanvir, Peter Laursen, Rohan P. Naidu, Charlotte A. Mason, Meghana Killi, Intae Jung, Tiger Yu-Yang Hsiao, Abdurro'uf, Dan Coe, Pablo Arrabal Haro, Steven L. Finkelstein, & Sune Toft.
Editor’s summary
Gas in galaxies provides the raw material for star formation. Galaxies in the early Universe are seen to be forming stars rapidly (see the Perspective by Scarlata), but the amount of gas they contain is difficult to determine observationally. Heintz et al. analyzed near-infrared spectroscopy of 12 galaxies at redshifts greater than eight, equivalent to less than 600 million years after the Big Bang. They identified three galaxies with characteristic rest-frame ultraviolet absorption caused by neutral hydrogen gas located in and around the galaxy. The high column densities of gas are sufficient to sustain the rapid star formation occurring in those galaxies, but only for a short period. —Keith T. Smith
Abstract
Primordial neutral atomic gas, mostly composed of hydrogen, is the raw material for star formation in galaxies. However, there are few direct constraints on the amount of neutral atomic hydrogen (HI) in galaxies at early cosmic times. We analyzed James Webb Space Telescope (JWST) near-infrared spectroscopy of distant galaxies, at redshifts ≳8. From a sample of 12 galaxies, we identified three that show strong damped Lyman-α absorption due to HI in their local surroundings. The galaxies are located at spectroscopic redshifts of 8.8, 10.2, and 11.4, corresponding to 400 to 600 million years after the Big Bang. They have HI column densities ≳1022 cm−2, which is an order of magnitude higher than expected for a fully neutral intergalactic medium, and constitute a gas-rich population of young star-forming galaxies.
Kasper E. Heintz et al.
Strong damped Lyman-α absorption in young star-forming galaxies at redshifts 9 to 11.
Science 384, 890-894 (2024). DOI:10.1126/science.adj0343
No wonder frauds like Ken Ham need to be so intellectually dishonest and teach their dupes to be at least as dishonest as they are, when scientists keep producing devastating evidence that the Bible is ludicrously wrong in its description of the creation of a little universe consisting of a small flat planet with a dome over it centred on the Middle East, just a few thousand years ago.
The age of the universe and the forces that produced it are such that the creation of galaxies scientists have now been there and witnessed for the first time, is exactly as science predicted from other strands of evidence than direct observation.
Advertisement
What Makes You So Special? From The Big Bang To You
How did you come to be here, now? This books takes you from the Big Bang to the evolution of modern humans and the history of human cultures, showing that science is an adventure of discovery and a source of limitless wonder, giving us richer and more rewarding appreciation of the phenomenal privilege of merely being alive and able to begin to understand it all.
Available in Hardcover, Paperback or ebook for Kindle
Advertisement
Ten Reasons To Lose Faith: And Why You Are Better Off Without It
This book explains why faith is a fallacy and serves no useful purpose other than providing an excuse for pretending to know things that are unknown. It also explains how losing faith liberates former sufferers from fear, delusion and the control of others, freeing them to see the world in a different light, to recognise the injustices that religions cause and to accept people for who they are, not which group they happened to be born in. A society based on atheist, Humanist principles would be a less divided, more inclusive, more peaceful society and one more appreciative of the one opportunity that life gives us to enjoy and wonder at the world we live in.
Available in Hardcover, Paperback or ebook for Kindle
This paper is enough to send a dedicated creationist disinformation specialist into intellectual summersaults. On the one hand, they hate the fact that dinosaurs lived so long ago and are evidence of life on earth tens and hundreds of millions of years ago, so they bend over backwards to prove they were around just a few thousand years ago, and then, horror of horrors, along come some clever scientists and show evidence that some dinosaurs even evolved into birds!
And now, courtesy of Palaeontologists at University College Cork (UCC) in Ireland , we have evidence of the transition from scales to feathers in pre-avian dinosaurs.
But it gets even worse! Creationists frauds have ben claiming for several years now that 'fossilised' soft dinosaur tissues prove they were just a few thousand years old because, despite the perfectly rational explanation of how 'soft' tissues can be preserved for many millions of years in the right circumstances, like the presence of iron, the evidence of this transition from scales to feathers would found in fossilised soft tissue!
So, a creationist disinformation specialist must now argue that fossilised dinosaur soft tissues don't exist, and when they do, they prove dinosaurs lived recently. One problem they face is that the fossilised skin isn't skin at all, or even the fibrous protein parts of skin; it has been entirely mineralised aa silicate, preserving only the morphology, including the surface patterning - which is where the evidence lies.
The University College Cork scientists have just published their findings, open access, in Nature Communications and in a UCC news release:
Genetics show the common origins of horned, hooved mammals
A diverse array of mammal headgear is on display in the Museum’s Richard Gilder Center for Science, Education, and Innovation as part of the Louis V. Gerstner, Jr. Collections Core.
Although the horns of cattle, gazelles and goats look very different to the antlers of deer and the ossicones on the head of a giraffe, and indeed, they are constructed differently, the cells they develop from in the embryo are the cells of the 'neural crest' that also develop into the face rather than the rest of the cranium. That and the fact that the underlying genetic control of their growth is sufficiently similar, provides compelling evidence that they share a common origin from which they, and the orders of which they are typical, have diverged.
This is the conclusion of two researchers at the American Museum of Natural History and Baruch College and the CUNY Graduate Center, who have just published their findings, open access in the journal, Communications Biology. It is also explained in an American Museum of Natural History press release:
Ducks, like almost all taxonomic categories, are a nightmare for creationists because they aren't mentioned in the Bible so it's not possible to put them into their primitive 'kind' classification, other than saying they are some 'kind' of bird, or maybe a 'duck kind'. The problem is there are so many different families of duck and several different species within each family. Some of them look very different to other ducks and have clearly evolved unique characteristics such as the long, serrated beak of the fish-eating mergansers, which places them in a different clade to the dabbling diving and torrent ducks.
By any definition then, evolving from a basic 'duck' into a merganser is the evolution of a new taxon, or 'macro-evolution' as creationists like to call it, while proclaiming it to be impossible because of some undefined magical process that intervenes to prevent it.
Which leaves creationists with nowhere to go but to claim mergansers were on the Ark, since they couldn't have evolved from a parent pair of 'ducks' at the abnormal warp-speed rate of evolution, that no-one noticed or thought to record, that creationists now claim must have happened in the few years since their favourite genocidal flood, that left no trace of itself.
But that's nothing compared to the genetic evidence that shows how, for example, the mergansers, evolved and spread geographically 18 million years before creationists believe there was a universe and again between 14 and 7 million years before creationists' god decided to create a small flat planet with a dome over it in the Middle East, 10,000 years ago, that creationists still believe accurately describes all that there is.
First, a little AI information about mergansers:
Tell me all about mergansers, their distribution and evolutionary origins, please. Mergansers are a group of ducks belonging to the genus Mergus, which is part of the family Anatidae. They are known for their distinctive serrated bills, which are well-adapted for catching fish, their primary diet. Here’s a detailed overview of mergansers, including their distribution, characteristics, and evolutionary background.
Characteristics
Mergansers are medium to large-sized ducks with slender bodies and long, pointed bills. Their bills are equipped with saw-like edges that help them grasp slippery fish. There are several species within the Mergus genus, including:
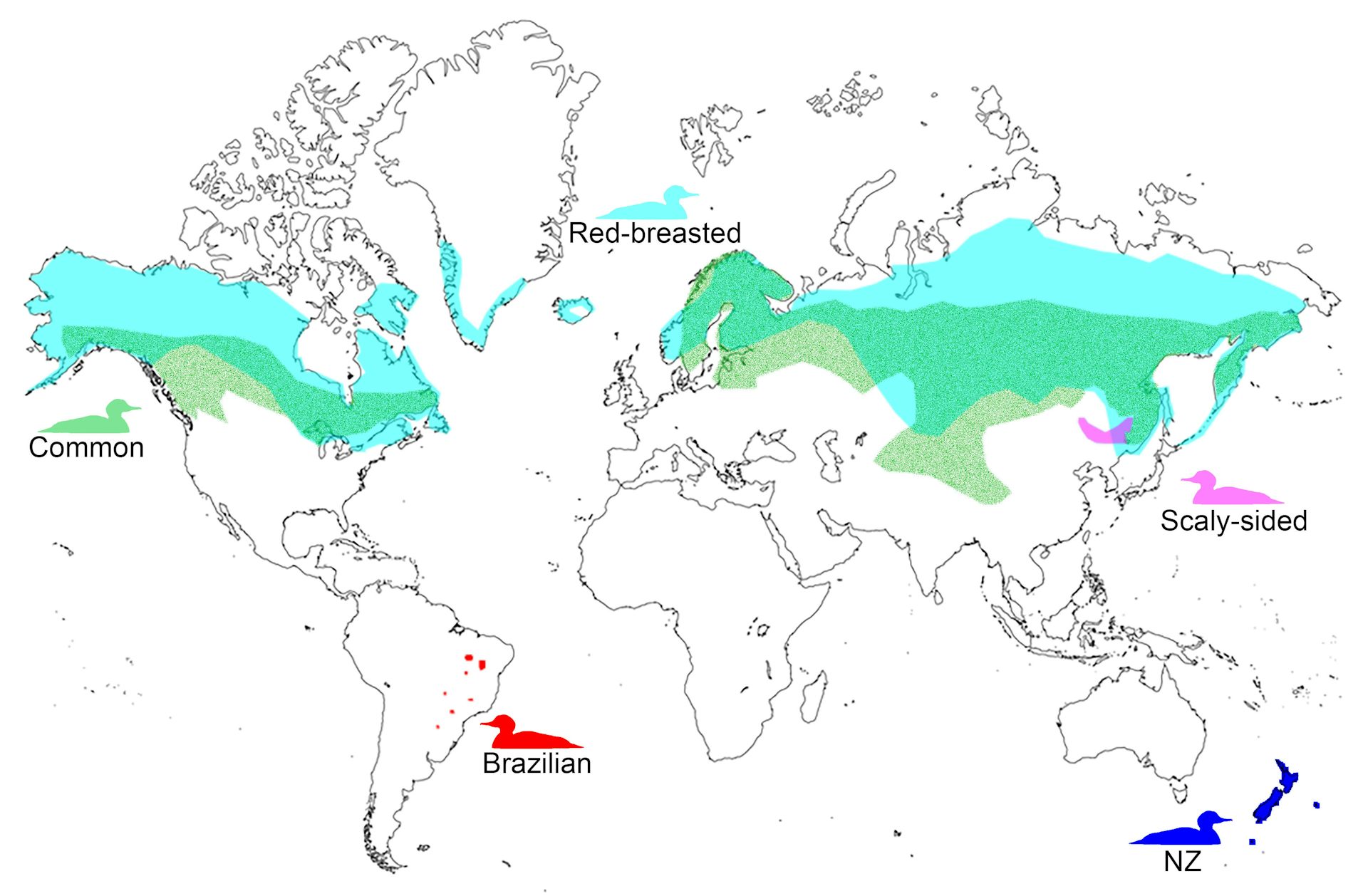
Common Merganser (Mergus merganser): Also known as the goosander, this species has a widespread distribution across North America, Europe, and Asia.
Red-breasted Merganser (Mergus serrator): Found in North America, Europe, and Asia, this species prefers coastal and large inland waters.
Hooded Merganser (Lophodytes cucullatus): Native to North America, it is smaller than other mergansers and has a distinctive fan-shaped crest.
Brazilian Merganser (Mergus octosetaceus): One of the rarest mergansers, it is found in central Brazil and is critically endangered.
Scaly-sided Merganser (Mergus squamatus): This species inhabits parts of East Asia and is also considered endangered.
Distribution
Mergansers have a wide distribution, with different species occupying various regions around the globe:
Common Merganser: Breeds in forested regions near lakes and rivers across northern North America, Europe, and Asia. In winter, they migrate to more temperate areas.
Red-breasted Merganser: Found in boreal and temperate regions across the Northern Hemisphere. They often winter in coastal waters, estuaries, and large inland lakes.
Hooded Merganser: Prefers forested wetlands, rivers, and ponds in North America. They are non-migratory in some parts of their range but move to ice-free waters in winter.
Brazilian Merganser: Restricted to central Brazil, particularly in the Serra da Canastra region. Their habitat includes clear, fast-flowing rivers.
Scaly-sided Merganser: Breeds in forested rivers of northeast China and the Russian Far East. They migrate to southern China, Korea, and Japan for winter.
Evolution
Mergansers belong to the Anatidae family, which includes ducks, geese, and swans. The evolution of mergansers is part of the broader adaptive radiation of the Anatidae, which diversified to exploit a variety of ecological niches. The serrated bills of mergansers are a specialized adaptation for piscivory (fish-eating), differentiating them from other ducks.
Evolutionary Background
Ancestral Origins: The Anatidae family likely originated in the late Cretaceous to early Paleogene periods. Early diversification allowed these birds to adapt to various ecological roles, including herbivory, omnivory, and piscivory.
Adaptive Radiation: The diversification of Mergus species is an example of adaptive radiation, where different species evolved to occupy different ecological niches, particularly in terms of habitat preference and diet specialization.
Morphological Adaptations: The distinctive serrated bills of mergansers are a key evolutionary trait that sets them apart from other ducks. This adaptation is crucial for their fish-based diet.
Conservation Status
Several species of mergansers are of conservation concern:
Brazilian Merganser: Critically endangered, with an estimated population of fewer than 250 individuals in the wild. Habitat destruction and water pollution are major threats.
Scaly-sided Merganser: Endangered, with threats including habitat loss due to logging and river damming, as well as pollution and human disturbance.
Conclusion
Mergansers are a fascinating group of ducks with specialized adaptations that enable them to thrive as piscivores in a variety of aquatic habitats. Their wide distribution and evolutionary history reflect the dynamic processes of diversification and adaptation in the Anatidae family. Conservation efforts are crucial for the survival of the endangered species within this group, highlighting the need for habitat protection and restoration.
The evidence of when the now extinct New Zealand merganser got to New Zealand and how it is related to other mergansers is the subject of an article in The Conversation by Nic Rawlence, Senior Lecturer in Ancient DNA, and Alexander Verry, Researcher, both of the University of Otago, New Zealand. Their article is reprinted here under a Creative Commons license, reformatted for stylistic consistency:
Ancient DNA from an extinct native duck reveals how far birds flew to make New Zealand home
Auckland Island merganser. Artistic reconstruction by J. G. Keulemans from Bullers Birds of New Zealand (1888)
Ask a bird lover if they have heard of the extinct giant moa or its ancient predator, Haast’s eagle, and the answer will likely be yes. The same can’t be said of New Zealand’s extinct, but equally unique, mergansers – a group of fish-eating ducks with a serrated bill.
The only southern hemisphere representatives of this group are the critically endangered Brazilian merganser and those from the New Zealand region, which are now extinct.
Unlike some of New Zealand’s other extinct birds, the biological heritage of our enigmatic mergansers is shrouded in mystery. But our new research on the extinct Auckland Island merganser is changing the way we think about the origins of New Zealand’s birds. Did the ancestors of the merganser come from South America or the northern hemisphere – and when did they arrive?
Mergansers are relatively common in the northern hemisphere but were limited to Brazil and the New Zealand region in the southern hemisphere.
Author provided
Lost to humans and pests
Mergansers were spread across the three main islands of New Zealand at the time of Polynesian arrival in the 13th century, as well as the Auckland Islands to the south and the Chatham Islands to the east.
Over-hunting, habitat destruction, and predation from the Pacific rat and Polynesian dog resulted in the extinction of mergansers on the New Zealand mainland and the Chatham Islands. By the time Europeans arrived in the 17th century, mergansers were restricted to an isolated population on the sub-Antarctic Auckland Islands.
European discovery of the Auckland Islands in 1806 led to a formal description of the Auckland Island merganser in 1841. However, European discovery brought new predators like pigs and cats.
Mergansers were also sought after as specimens for the museum trade. The last known Auckland Island merganser was shot and collected in 1902, a mere 61 years after its discovery.
Auckland Island Merganser, Mergus australis, collected June 1902, Auckland Islands, New Zealand.
Only recently have merganser bones from the Chatham Islands been described as a distinct species. The distinction of the Chatham Island merganser hints at greater merganser diversity in the New Zealand region than previously thought. It is possible the merganser bones found on mainland New Zealand may be another species, but only ongoing research will be able to confirm this.
From the northern hemisphere to NZ
The extinction of mergansers from the New Zealand region has meant their evolutionary history has remained a mystery. Did their ancestors, and those of the Brazilian merganser, arrive via independent colonisation events from the northern hemisphere? Or was there a single push into the southern hemisphere, followed by subsequent divergence events?
To find out more, we sequenced ancient DNA from an Auckland Islands merganser and a Brazilian merganser. This allowed us to reconstruct the evolutionary history of the wider group.
We found mergansers originated in the northern hemisphere, diverging from their closest relatives some 18 million years ago, before rapidly evolving into several different species between 14 and seven million years ago.
The mergansers from the New Zealand region are most closely related to the northern hemisphere common merganser. Their ancestors arrived here at least seven million years ago in a separate colonisation event to the one that gave rise to the Brazilian merganser.
Further genetic research is currently underway. The goal is to reconstruct the evolutionary history of mergansers within the New Zealand region.
The global origins of New Zealand’s birds
Many New Zealanders believe the country’s native birds originate from Australia. Increasingly though, genetic and palaeontological research shows a number of our feathered friends hail from further afield.
The long journey of blue-eyed shags started in South America, with the birds island hopping via Antarctica and the sub-Antarctic islands to New Zealand. Mergansers arriving from the northern hemisphere add another piece to the puzzle.
It is possible that fossils of extinct mergansers (and other birds with distant geographic origins) will be discovered as palaeontologists increasingly focus on previously neglected and newly discovered southern hemisphere fossil deposits.
Only then, combined with the power of ancient DNA, will we be able to fully understand how New Zealand’s dynamic geological, climatic and human history has influenced the colonisation and diversification of birds on this isolated South Pacific archipelago. Nic Rawlence, Senior Lecturer in Ancient DNA, University of Otago and Alexander Verry, Researcher, Department of Zoology, University of Otago
Abstract
Mergansers are riverine and coastal piscivorous ducks that are widespread throughout North America and Eurasia but uncommon in the Southern Hemisphere. One species occurs in South America and at least two extinct species are known from New Zealand. It has been proposed that these Southern Hemisphere merganser lineages were founded by at least two independent dispersal events from the Northern Hemisphere. However, some morphological and behavioural evidence suggests that Southern Hemisphere mergansers may form a monophyletic clade that descended from only a single dispersal event from the Northern Hemisphere. For example, Southern Hemisphere mergansers share several characteristics that differ from Northern Hemisphere mergansers (e.g. non-migratory vs. migratory, sexual monochromatism vs. sexual dichromatism, long vs. short pair bonds). We sequenced complete mitogenomes from the Brazilian merganser and an extinct merganser from New Zealand—the Auckland Island merganser. Our results show that the Brazilian and Auckland Island mergansers are not sister-taxa, and probably descend from two separate colonization events from the Northern Hemisphere at least 7 Mya. Nuclear (palaeo)genomic data may help to further resolve the relationship between living and extinct mergansers, including merganser fossils from New Zealand that have not been subjected to palaeogenetic analysis.
INTRODUCTION
Mergansers (Mergus spp.) are a group of riverine and seasonally coastal fish-eating ducks that have a widespread Northern Hemisphere distribution but are uncommon in the Southern Hemisphere (Kear 2005, Williams et al. 2012, 2014) (Fig. 1). They are characterized by a serrated bill, and include the endangered scaly-sided merganser (M. squamatus Gould 1864) from north-east Asia; the common merganser (M. merganser Linnaeus 1758), and the red-breasted merganser (M. serrator Linnaeus 1758), which have widespread Northern Hemisphere distributions; the critically endangered Brazilian merganser (M. octosetaceus Vieillot 1817); and two currently recognized extinct species from the New Zealand region—M. australis Hombron and Jacquinot 1841 and M. milleneri Williams and Tennyson 2014 from the Auckland and Chatham Islands, respectively. While the hooded merganser Lophodytes cucullatus (Linnaeus 1758), previously M. cucullatus, from North America has a serrated bill, it is not considered a ‘true’ merganser (e.g. Buckner et al. 2018, Lavretsky et al. 2021). The taxonomic relationship of the smew Mergellus albellus (Linnaeus 1758) from Eurasia is currently unresolved; it is sometimes suggested to be more closely related to Mergus and Lophodytes or to goldeneyes (Bucephala spp.) (Livezey 1995, Buckner et al. 2018, Lavretsky et al. 2021).
Figure 1.
Schematic of the breeding distributions of Mergus spp. The New Zealand (NZ) lineage encompasses the Auckland Island merganser (465 km south of NZ) and Chatham Island merganser (785 km east of NZ), as well as Mergus spp. from mainland NZ. Breeding distributions are based off the Cornell Lab of Ornithology Birds of the World website.
The now extinct Auckland Island merganser M. australis (or miuweka) (Fig. 2) was formally described in 1841, based on a specimen collected on the sub-Antarctic Auckland Islands, 465 km south of mainland New Zealand. Rare Late Holocene-aged merganser bones have been found in coastal sand dune deposits (including Māori middens) on New Zealand’s three main islands (Stewart, North, and South), and the Auckland and Chatham Islands (Tennyson and Martinson 2007, Williams et al. 2014, Tennyson 2020). Bones from the latter were recently described as a distinct species M. milleneri, which was smaller than the nominate M. australis, with a shorter skull, relatively shorter premaxilla, smaller sternum and keel, relatively shorter wing bones, and a narrower pelvis (Williams et al. 2014). The taxonomic status of merganser bones from mainland New Zealand is unresolved (i.e. cannot be assigned to either M. australis or M. milleneri), and are currently recognized as Mergus spp. (Birds New Zealand Checklist Committee 2022).
Figure 2.
In the Southern Hemisphere, mergansers are only known from the New Zealand region and South America, represented here by the Auckland Island merganser. A, artistic reconstruction by J.G. Keulemans from Buller (1888); B, historical museum skin (Museum of New Zealand Te Papa Tongarewa OR.001357); C, the Brazilian merganser (photo by Savio Freire Bruno CC BY-SA 3.0).
Mergansers in the New Zealand region are mainly thought to have occupied a riverine and seasonally coastal habitat (e.g. sheltered bays during winter; Kear 2005, Williams et al. 2012, 2014). It is likely that they mainly nested in tree cavities, but also caves in some instances, as the remains of adults, chicks, and eggs have been found within a cave on the Chatham Islands (Williams et al. 2014). By the 17th century, mergansers had been extirpated from the Chatham Islands and mainland New Zealand, and survived only on the Auckland Islands. A combination of subsistence hunting, and predation from the Pacific rat (Rattus exulans) and Polynesian dog (Canis familiaris), probably led to the extinction of mergansers across most of their prehistoric range (Tennyson and Martinson 2007, Greig and Rawlence 2021.1). On the Auckland Islands, predation from introduced pigs (Sus scrofa) and cats (Felis catus), and collecting for the museum trade, resulted in their extinction—indeed the last known Auckland Island merganser specimen was shot and collected in January 1902 (Williams 2012.1).
The only extant merganser in the Southern Hemisphere—the critically endangered Brazilian merganser (Fig. 2)—is one of the rarest birds in the world, comprising only 250 wild individuals. It is split across three remnant populations in Brazil, but once had a more widespread historical distribution encompassing Argentina and Paraguay (Vilaca et al. 2012.2, Maia et al. 2020.1). The Brazilian merganser has undergone a significant population bottleneck, yet different remnant populations can still be genetically identified (Maia et al. 2020.1). Like mergansers from the New Zealand region, the Brazilian merganser occupies riverine habitats, and often nests in tree cavities or rock crevasses (Vilaca et al. 2012.2, Maia et al. 2020.1).
It has been proposed that the Southern Hemisphere mergansers were founded by independent dispersal events to the New Zealand region and South America from the Northern Hemisphere (e.g. Livezey 1995). Based on behavioural characteristics, Johnsgard (1961) tentatively assigned the Brazilian merganser as sister-species to a clade comprising the other Mergus species, with the Auckland Island merganser as the sister-species of the common merganser and scaly-sided merganser. In contrast, using morphological characters, Livezey (1989, 1995) assigned the Auckland Island merganser, then Brazilian merganser, as successive sister-species to all other Mergus species, though with weak to moderate bootstrap support. Using mitochondrial DNA (mtDNA) sequences, Buckner et al. (2018) suggested the Brazilian merganser was the sister-species to the scaly-sided merganser, albeit with weak support. However, some evidence suggests that Southern Hemisphere mergansers may be closely related to one another, potentially even sister-species, as they share several behavioural (e.g. non-migratory and long pair bonds) and morphological (e.g. sexually monochromatic) characteristics, in contrast to their Northern Hemisphere congeners (e.g. migratory, short pair bonds, and sexual dichromatism; Livezey 1995). In addition, recent genetic studies of other extinct Southern Hemisphere avian species have also revealed unexpected evolutionary connections between birds from New Zealand, South America, and Africa (e.g. Mitchell et al. 2014.1a, 2014.2b, Boast et al. 2019, Rawlence et al. 2022.1, Verry et al. 2022.2a). As such, the phylogenetic relationships of the Southern Hemisphere mergansers, when their ancestors arrived in the region, and from where, remain unresolved.
In this study, the first genetic study of a New Zealand Mergus species, we sequenced mitochondrial genomes (mitogenomes) from historical museum specimens from the Auckland Island merganser and Brazilian merganser, and analysed them within a phylogenetic framework of Mergini mitogenomes (Liu et al. 2012.3, Lavretsky et al. 2021). These data were used to determine the phylogenetic relationships and divergence dates within mergansers.
Creationists needn't be too embarrassed by these facts about the distribution and evolution of mergansers, after all, the simple Bronze Age Canaanites who made up the origin myths to fill the gaps in their knowledge and understanding, could have known nothing of them, since they are largely absent from the Middle East, nor of southern hemisphere land masses such as South America, Australia and New Zealand when they didn't even realise there is a southern hemisphere, or of the ducks and their families.
And, of course, they knew nothing about the history of Earth and the life on it, so how could they be expected to have known about the evolutionary history and geographic distribution of these ducks? With their long, serrated beaks and fish diet, they probably wouldn't have recognised mergansers as a type of duck even.
All they had to draw on was their limited knowledge of the small scrap of the globe they inhabited, which is why they got almost everything they described so hopelessly wrong. The lesson from Genesis is just how badly you do when you know little and depend on guesswork from the point of view of someone with the knowledge understanding of a pre-school toddler of today.

 #
#