Reconstruction of Musango matusadonaensis in its natural setting
AI-generated image (ChatGPT 5.6 Sol)
Based on an illustration by Mark Witton.
Based on an illustration by Mark Witton.


When Musango matusadonaensis was alive, it would have lived alongside ancient rivers and streams.
© Mark Witton 2026
It is gap-filling time again — not because palaeontologists have discovered some predetermined evolutionary “missing link”, but because another conspicuous blank on the palaeontological map is being filled. This time, the new evidence comes from Africa and concerns a dinosaur that lived about 210 million years ago, during the Late Triassic.
The discovery sheds new light on the early diversification and geographical distribution of dinosaurs, when they were still only a relatively minor component of terrestrial ecosystems. Over the following millions of years, they diversified into an extraordinary range of forms and became the dominant large land animals of the Jurassic and Cretaceous periods.
Gaps in the fossil record are, of course, eagerly seized upon by creationists desperate to find somewhere to insert their god. Their enthusiasm tends to diminish markedly whenever palaeontologists recover new evidence from one of those gaps. Each such discovery removes another supposed refuge for the ever-shrinking god of the gaps, so the evidence must be dismissed as misidentified, incorrectly dated or merely another example of “evolutionary storytelling”.
No matter how many gaps science fills, creationists continue to assure themselves that the next one will be different: that it will finally be the gap that science cannot investigate and into which their preferred creator can safely be inserted. Unfortunately for that strategy, palaeontologists continue to discover fossils, dating methods continue to improve, and previously neglected regions continue to be explored.
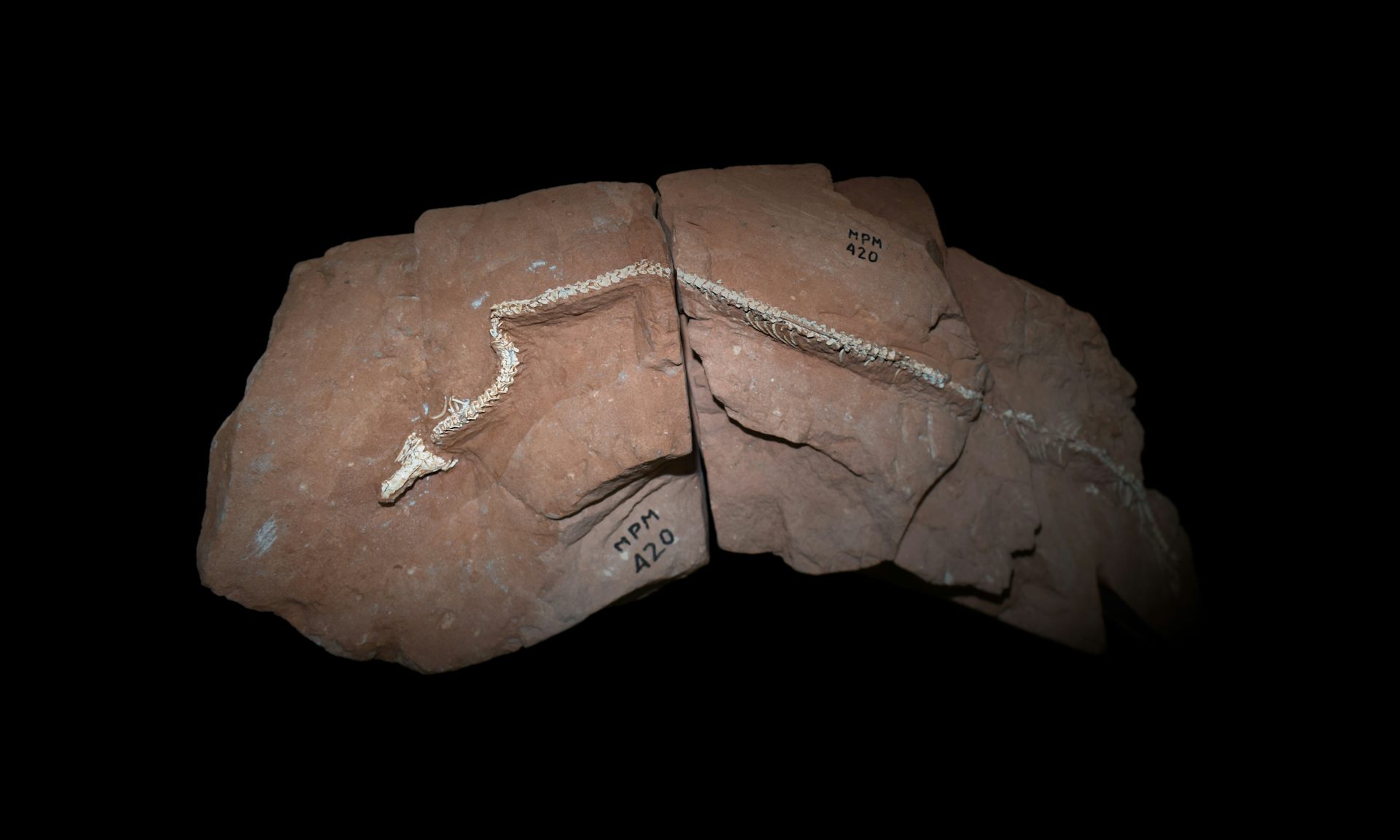
This latest discovery consists of a partial skeleton excavated from exposures of the Late Triassic Pebbly Arkose Formation on the shore of Lake Kariba in northern Zimbabwe. An international team of Zimbabwean, South African and British scientists has named the animal Musango matusadonaensis. The study was led by Professor Paul M. Barrett of the Natural History Museum, London, while the expeditions during which the fossils were collected were led by Professor Jonah Choiniere of the University of the Witwatersrand’s Evolutionary Studies Institute.
Musango was an early sauropodomorph — a member of the lineage that later produced the enormous, long-necked sauropods such as Diplodocus. Unlike those gigantic four-legged descendants, however, Musango was a relatively lightly built, predominantly bipedal animal. It measured an estimated 4.5 metres in length but weighed only about 222 kilograms. Its skull has not been recovered, so its diet cannot be established with certainty, although comparisons with related dinosaurs suggest that it was probably herbivorous or omnivorous.
The fossil record of early dinosaurs is extremely uneven. Europe, North America and parts of South America have been studied far more intensively than much of Africa, leaving large gaps caused not necessarily by the absence of dinosaurs but by the scarcity of sustained palaeontological exploration. Musango is only the fifth dinosaur formally named from Zimbabwe, following the description in 2024 of another sauropodomorph from the same formation, Musankwa sanyatiensis.
Phylogenetic analysis places Musango and Musankwa as close relatives within Unaysauridae, a group whose fossils are known from Africa, India and South America — landmasses that were then joined within Gondwana. Together, these discoveries support an emerging picture in which Late Triassic southern Africa contained several distinct ecosystems with their own dinosaur communities, rather than a single uniform fauna extending across the region. Africa was therefore not a peripheral spectator in early dinosaur evolution; its still-underexplored rocks may preserve crucial evidence about how dinosaurs diversified and spread before their later ecological dominance.