Creationists will continually demand evidence for evolution being observed, then, when the evidence is provided, immediately insist that science should adopt their childish parody of evolution, in which one species turns into an unrelated species in a single miraculous event. That is not evolution as any biologist understands it. In fact, if such a thing were ever observed, it would falsify the Theory of Evolution, not confirm it.
By demanding evidence for something no scientist has ever claimed happens, creationists imagine they are somehow refuting science, or at least providing a plausible anti-Darwin argument for people who do not understand the science.
So this example of evolution in living populations will almost inevitably be dismissed by creationists using that same disingenuous tactic. It is evidence for the evolution of island gigantism in isolated populations of the wren, Troglodytes troglodytes, on Scottish islands. And, to rub salt in creationists' wounds, it is not merely a single isolated example, but multiple examples of gigantism evolving in island environments — an example of parallel evolution in response to similar environmental pressures acting on different local populations.
In other words, this is not some local curiosity that can be waved away as a one-off oddity, but the predictable result of isolation, restricted gene flow and similar island conditions acting on related populations. The evidence has just been published, open access, in the Evolutionary Journal of the Linnean Society by researchers led by the University of Birmingham.
The researchers, led by Dr Michał Jezierski, examined four subspecies of island wren, each isolated on a specific Scottish island or archipelago — Shetland, Fair Isle, the Outer Hebrides and St Kilda. Each of these subspecies is geographically isolated, yet exposed to broadly similar island environments, and each differs significantly from the wrens found throughout mainland Britain and continental Europe.
The study showed that the wrens of St Kilda and Shetland show little evidence of interbreeding with the mainland population. These two populations have evolved spectacular island gigantism: a wren from England will typically weigh about 7–10 grams, while a St Kilda wren weighs about 13–16 grams. The largest St Kilda wrens are therefore more than twice the weight of the smallest mainland wrens, and their genetic distinctiveness is so marked that the researchers say they may be on the way to becoming separate species.
Importantly, the genomic evidence shows that the Shetland and St Kilda wrens are genetically distinct from each other, despite having evolved similar enlarged body sizes. In other words, the same broad evolutionary outcome has arisen independently in separate island populations, rather than being inherited from a single already-giant ancestor. That is exactly what evolutionary biology predicts: related populations, isolated in similar environments, can be shaped in similar directions by similar selection pressures, even when the detailed genetic route differs.
Because many creationists never seem to have progressed beyond the teleological thinking typical of toddlers, they assume that everything must have agency and must be directed towards some intended end.
That habit of mind was highlighted a few years ago in a study showing that the persistence of teleological thinking into adulthood is associated with the cognitive style underlying both creationism and conspiracism.
This goes a long way towards explaining why creationists so often invoke conspiracies to explain away the evidence against their beliefs, and why debating them can resemble arguing with toddlers who know little, understand less, and yet imagine themselves in possession of profound truths.
Any serious study of evolution, however, quickly shows that there is no plan, no foresight, and no directing intelligence. Evolution has no destination in mind. The only “direction” it has is whatever changing environments impose on it, favouring traits that happen, at any given time, to work best and leave the most descendants.
A recent study by a team led by Dr Yosef Kiat of the School of Zoology at Tel Aviv University’s George S. Wise Faculty of Life Sciences and the Steinhardt Museum of Natural History, working with colleagues in China and the United States, provides a striking illustration of that point. Their paper, published on 21 November 2025 in Communications Biology, suggests that some feathered dinosaurs in the anchiornithid lineage of pennaraptoran theropods may have evolved the ability to fly, only to lose it again when environmental change made flight no longer advantageous.
In other words, evolution did not move steadily towards some grand objective. It produced flight when flight was useful, and abandoned it when it was not. Feathers, which may briefly have served an aerodynamic role in these animals, appear then to have reverted primarily to their earlier functions of insulation and temperature regulation.
The key evidence comes from moulting patterns. In living flying birds, feathers are moulted symmetrically so that the bird can retain its ability to fly. In flightless birds, by contrast, moulting can be irregular and apparently disorganised. By examining fossils of these dinosaurs from Chinese deposits, the researchers identified this latter pattern in animals that otherwise appear to have possessed anatomical adaptations for flight.
A male satin bowerbird by his highly decorated avenue bower.
Photo by Daniel J. Field
Tiny fossil bone helps unlock history of the bowerbird | University of Otago
Apart from the fact that this fossil is a million years old, there is nothing in this discovery that creationists will struggle to dismiss with one of their well-worn stock phrases — “It was just a bird ‘kind’,” “It wasn’t transitional,” and so on. This is despite the fact that their Bible is remarkably vague about how many bird ‘kinds’ there were, includes bats as birds, and says absolutely nothing about anything outside a few square miles of the Middle East.
And of course, the date — like the entire fossil record — will be casually brushed aside as forged, fabricated, or “wrongly dated using proven false carbon dating” [sic].
But to anyone who actually values evidence and truth, and is not intent on proving their strength by clinging to demonstrably false beliefs in defiance of all contrary evidence, this find is genuinely fascinating. It provides strong evidence that the bowerbirds, today confined to Australia and New Guinea, were once far more widespread. This conclusion is based on the fact that the fossil was discovered in New Zealand. It is also suggested that climate change may have brought about its extinction in New Zealand and driven the bowerbirds' range back to its present distribution.
The discovery is reported in the journal Historical Biology by researchers from University of Cambridge, University of Otago, and Museum of New Zealand Te Papa Tongarewa. A [news release from the University of Otago]() explains the significance of the find and four of the authors have also written an article about the find in The Conversation. Their article is reprinted here under a Creative Commons licence, reformatted for stylistic consistency.
A recent paper published in Nature Ecology & Evolution lends support to Darwin’s theory that language evolved from learned responses to innate sounds conveying specific information — such as a scream in response to pain — behaviours shared across many species.
The paper, by a team of researchers led by William E. Feeney of the Doñana Biological Station, Spanish National Research Council (CSIC), Seville, Spain, and the School of Environment and Science, Griffith University, Nathan, Queensland, Australia, together with James A. Kennerley of the Cornell Lab of Ornithology, Ithaca, New York, USA, suggests that different bird species—separated by thousands of miles geographically and tens of millions of years evolutionarily—may illustrate Darwin’s theory in practice.
This study not only strengthens Darwin’s insight into the evolutionary roots of human language but also highlights how universal biological processes—rather than supernatural design—can account for the complexity of communication across species.
Feeney and Kennerley, with co-author Niki Teunissen of Monash University, have explained their discovery and its significance in an article in The Conversation, reprinted here under a Creative Commons licence and reformatted for stylistic consistency.
(a) Blue Jay by Travis Maher (ML578309451). Cornell Lab of Ornithology, Macaulay Library.
(b) Hybrid Jay by Brian R. Stokes.
(c) Green Jay by Dan O'Brien (ML390361871). Cornell Lab of Ornithology, Macaulay Library.
An interesting example of hybridisation between two distantly related species of jay has been reported by two biologists at the University of Texas, Austin. The hybrid is a cross between a blue jay (Cyanocitta cristata) and a green jay (Cyanocorax yncas). Although both are jays — members of the crow (Corvid) family — these two belong to different genera that diverged some seven million years ago and, until recently, lived in non-overlapping ranges in North America. Climate change has allowed them to extend their ranges, bringing them into contact only within the past decade.
The reason this is scientifically significant is that it illustrates how divergent evolution proceeds, and why it does not always progress to complete reproductive isolation. Unlike many other vertebrates, the genomic arrangements of birds are remarkably stable. As long as alleles remain in the same chromosomal loci, interbreeding remains possible. This can be disadvantageous if it results in hybrids with reduced fitness. For example, one population may evolve a long, slender bill adapted for one type of seed, while another evolves a short, stout bill for harder seeds. A hybrid might inherit an intermediate bill suited to neither, creating obvious disadvantages. In such cases, natural selection favours the evolution of barriers to hybridisation.
In many organisms, this is achieved by genetic rearrangements that prevent a zygote from developing even when mating occurs—so-called post-zygotic barriers. Where genomes are stable, such rearrangements rarely arise, so species tend instead to evolve pre-zygotic barriers that prevent mating or fertilisation in the first place. Among birds, these often take the form of plumage differences, mating rituals, or song—hence their remarkable diversity.
The Balearic shearwater (Puffinus mauretanicus, above) and the Mediterranean shearwater (Puffinus yelkouan, below) are two shearwater taxa that are genetically similar despite displaying morphological and migratory differences. The new study shows that these two taxa have undergone recurrent episodes of divergence and hybridization during the Pleistocene glacial and interglacial cycles, respectively. This serves as a key example of how hybridization can help preserve genetic diversity and evolutionary potential in threatened taxa.
Like the story of human evolution, that of two seabirds in the shearwater family living in the Mediterranean shows a history of divergence, followed by episodes of hybridization and genetic remixing. Normally, from a conservation perspective, hybridization is considered a problem because it can erode the distinctiveness of threatened species, diluting unique traits.
However, in the case of the Mediterranean shearwater (Puffinus yelkouan) and the critically endangered Balearic shearwater (Puffinus mauretanicus), hybridization has had the opposite effect. By introducing genetic diversity into the Balearic shearwater, it may have improved the species’ chances of survival, potentially making the difference between persistence and extinction.
This evolutionary story sits uneasily with the worldview of Bible-literalist creationists. The very fact of species interbreeding undermines the notion of fixed, separately created “kinds.” It also contradicts the idea that a few thousand years ago all life was destroyed in a global flood, followed by a period of hyper-rapid diversification into the species we see today—a process which, according to creationist claims, left no trace in the fossil record. If hybridization had occurred during that supposed burst of post-Flood diversification, it would simply have blurred the lines between “kinds,” creating variation within species rather than producing the multitude of distinct species we observe today. In other words, the evidence shows that life’s history is one of gradual divergence, occasional genetic exchange, and long-term adaptation—not sudden, discrete acts of “special creation.”
A recent study led by Professors Julio Rozas, Marta Riutort, and Jacob González-Solís of the Faculty of Biology and the Biodiversity Research Institute of the University of Barcelona (IRBio), together with Joan Ferrer Obiol of the University of Milan, has revealed how this evolutionary pattern unfolded. Using genomic analysis, they showed that the two shearwater species have gone through repeated cycles of divergence and hybridization, largely driven by the glacial–interglacial rhythms of the Pleistocene. Each cycle of isolation and secondary contact left genetic signatures that are still visible today.
The researchers found a clear gradient of genetic differentiation across the Mediterranean, with hybrid populations becoming increasingly common as one moves westwards, culminating in the Balearic Islands of Ibiza and Formentera. Intriguingly, the shearwater population on Menorca is genetically closer to P. yelkouan than to P. mauretanicus, reflecting its history of admixture. Far from being detrimental, this interbreeding has enriched the Balearic shearwater’s genome, reducing inbreeding risks and helping to preserve adaptive traits that may otherwise have been lost.
Scientists never set out to prove that the Biblical account of science and history as related in Genesis is hopelessly wrong and based on the childish guesses of scientifically illiterate people—that is simply an incidental outcome of the facts they revealed. Nevertheless, it remains a fact. The Bible presents a timeline and an account of the origin of species that are wholly inconsistent with the known facts.
To be fair to the original authors, they probably never intended to mislead scientifically illiterate people thousands of years later. As they devised origin myths to fill the gaps in their knowledge of the world, they could hardly have imagined that someone would one day write their tales down, combine them with other implausible myths, genealogies, and morality stories designed to spread fear and enhance priestly power, and then declare the compilation to be the inerrant word of an omniscient creator god. That declaration, of course, reinforced the text’s usefulness as a source of excuses for actions such as land theft, genocide, and enslavement—atrocities conveniently blamed on a god to absolve perpetrators of personal responsibility.
Remote islands are the sort of environment biologists might dream up if asked to design a natural laboratory for testing evolution. It’s no coincidence that Darwin was inspired to develop his theory while visiting the Galápagos Islands and noticing how the finches had adapted in different ways to the conditions on each island.
Another striking example comes from the Rēkohu Chatham Islands, about 785 kilometres east of mainland Aotearoa New Zealand. The islands rose in their present form around 3.5 million years ago, effectively resetting the clock for the ecosystems that would develop there. Species arriving from elsewhere had to make do with what traits they already carried, and only those suited to island life survived. Most new arrivals were birds, insects, or wind-blown plants carried there by chance. With few predators and limited competition, these colonists had the perfect opportunity to go their own evolutionary way.
Information about endogenous retroviruses is normally unwelcome news for creationists because they form phylogenies which exactly map onto the evolution of different species from common ancestry. This is no less true of a new research paper published by four researchers from the College of Life Sciences, Nanjing Normal University, Nanjing, Jiangsu, China, published in Molecular Biology & Evolution.
Endogenous retroviruses for part of the 'junk' (non-coding/non-regulatory) DNA, but some of it have been exapted for other functions over the years and some of it has placed an evolving taxon onto a new evolutionary trajectory. For example, one exapted retrovirus with immuno-suppressive qualities has made placental mammals possible without the growing embryo being treated as a parasite and attacked by the mother's immune system.
The researchers have uncovered the complex evolutionary history of retroviruses in birds by analysing their genetic “fossils” — endogenous retroviruses (ERVs) — embedded in bird genomes. They scanned the genomes of 758 bird species and identified more than 470,000 ERV sequences, revealing a vast and previously underestimated diversity of retroviruses. These sequences are the remnants of ancient viral infections that became part of the host DNA, passed down from generation to generation.
Feathers provide a fascinating example of how evolution can repurpose structures over time. Initially evolving in response to one set of selective pressures, feathers later opened the door for entirely new functions unrelated to their original purpose.
Early feathers appeared among dinosaurs primarily as an adaptation for thermoregulation. Simple, filamentous feathers offered significantly better insulation than traditional reptilian scales, helping dinosaurs maintain stable body temperatures. Among bipedal theropod dinosaurs, these insulating feathers eventually evolved into more complex structures, freeing forelimbs to develop into wings. Feathers subsequently became specialized for powered flight, having first likely served intermediate functions such as display or gliding.
The presence of insulating feathers likely provided a survival advantage during the dramatic climate changes following the asteroid impact that marked the end of the Cretaceous period, approximately 66 million years ago, contributing to the extinction of non-avian dinosaurs.
This is why we see a clear progression through the fossil record, unlike what would be expected of an intelligent design process, where birds, complete with flight feathers, would be expected to make a sudden appearance without ancestry.
Today, feathers in modern birds retain their important role in thermoregulation while also facilitating flight and serving as display structures. Vibrant and diverse plumage has evolved under sexual selection pressures, playing a crucial role in mating rituals, mate choice, and reinforcing genetic isolation among closely related species, thereby preventing hybridization.
As though designing birds with wings that they can't fly with wasn't stupid enough, it seems creationism's idiot designer designed them with flight feathers too. That is, if you believe the childish nonsense of special creation of species without ancestors, which is a central superstition of the creationist cult. And presumably, because it's also central to the cult that species don't evolve, it must be assumed that every extinct species was created without ancestors too, so they can't have evolved from ancestors either.
Which makes it all the more puzzling that a study has shown that as flightless birds became flightless over time, they tended to retain feathers that were characteristic of flying birds, and in particular, those of their flying relatives.
In other words, as flightlessness evolved, the last thing to change were their feathers.
Which begs thew question why creationism’s putative designer gave flightless birds feathers that looked as though they had been inherited from flying ancestors.
Of course, there is a rational explanation for this, and it doesn't involve magic creation by a blundering idiot behaving like a mindless process operating without a plan.
Hirundo rustica rustica (left) and Hirundo rustica tytleri (right) have different sexually selected traits. For example, one has a red chest and the other has a pale chest.
This paper in Science passed me by last December but it's too good to ignore because it is an example of something that creationists insist has never been seen because it doesn't happen - evolution in progress. And, to annoy creationists even more, it's an example of something Charles Darwin proposed as a mechanism for speciation - mate selection - as the isolating mechanism.
Barn swallows are one of the species that has formed a commensal relationship with humans that does not, like many other commensal species, involve it in consuming our crops either in the fields or in storage, from when we began building permanent or semi-permanent buildings. They are now dependent on our barns and out-houses as nesting places.
Moult remains of feather-feeding beetle larvae intimately associated with downy feather portions from an unidentified theropod dinosaur in Early Cretaceous amber of Spain. Insets show the head with powerful mandibles of one of the larval moults (top) and the pigmentation pattern of feather second order branches (bottom), with the main stem of one feather at the right of the amber fragment. The length of the amber fragment is 6 millimetres across.
Isolated moult of the feather-feeding beetle larva found in the Spanish amber outcrop of Rábago/El Soplao, with detail of its powerful mandibles (right). Length of the moult is less than two millimetres.
Image credit: CN IGME-CSIC.
To normal people, the discovery of feather-eating beetles and feather fragments in 105-million-year-old amber, 30 million years before there were birds might be a clue that something had feathers before bird had them, suggesting that birds might have evolved from whatever that was.
Not so, creationists, however. Creationists conclude that any evidence that doesn't agree with them must be wrong because their evidence-free dogma is sacred and therefore uninfluenced by real-world evidence.
So, the following is just something else for creationists to ignore while they pretend to know better than the experts who have, unlike creationists, actually studied the subject.
It is news that a study, co-led between the Geological and Mining Institute of Spain of the Spanish National Research Council (CN IGME-CSIC) and Oxford University Museum of Natural History (OUMNH) has shown that beetles fed on the feathers of dinosaurs about 105 million years ago. This is based on an analysis of spectacular fossil amber fragments, from the locality of San Just in north-eastern Spain, revealed moults of tiny beetle larvae tightly surrounded by portions of downy feathers.
The feathers belonged to an unknown theropod dinosaur that lived around 105 million years ago, during the Early Cretaceous. This means that the feathers could not have come from a ‘modern bird’ species, since current evidence indicates that this group appeared about 30 million years later in the fossil record, during the Late Cretaceous.
Unlike an intelligent designer, the process of evolution can't go back to basic and start again. It is normally an additive process that has no control over what it has to work with and simply refines and improves on what is there. That's not to say new structures can't evolve but they do so by enlarging or remodelling something that was already there - the membrane of a bat's wing, for example is the webbing that exists in the tetrapod embryo between the fingers and toes, while the feathers of a bird's wing are highly modified scales. Both those structures evolved out of tissues that were already there. It would have been impossible for a bat to grow wing feathers instead of a membrane, for example, because the earliest mammals had lost their scales and evolved fur.
But of course, that would not have been a problem for an omnipotent intelligent designer who, having designed one wing would not need to set about designing another way to do the same thing.
So, constrained as evolution was by what it could use, it's not really surprising that birds and bats evolved on two different trajectories, with the only thing in common being flight (and of course the basic vertebrate skeletal body plan).
The Broad-billed Tody, Todus subulatus, is a member of the bird group Coraciimorphae.
Image credit: Daniel Field, University of Cambridge
The Northern Flicker, Colaptes auratus, is a member of the bird group Coraciimorphae. Berv and co-authors identify this group of birds and others as having close ties to the end-Cretaceous mass extinction that occurred in the wake of the Chicxulub asteroid impact approximately 66 million years ago.
Image credit: Daniel Field, University of Cambridge
Another major milestone in the history of life on Earth happened, like almost everything else, in that long pre-Creation Week history that creationists need to ignore. It was the mass extinction about 66 million years ago that killed the non-avian dinosaurs and most megafauna, leaving vacant niches that could be exploited by the descendants of survivors. It's no surprise to anyone who understands how evolution works, that this led to a proliferation of new species as existing species diversified to fill those niches.
That much was known already from the fossil record, but now a University of Michigan study has found how this maps onto changes in the genome of the major bird families, as a kind of DNA fossil, just as the TOE predicts.
The major difference between the different taxons is how developed the chicks are on hatching and how dependent they are on their parents. In ducks, geese, the ratites and ground-nesting birds such as the plovers, chickens and turkeys, the chicks are mobile and able to feed themselves (precocial) almost as soon as they've hatched. At the other end of the scale, many passerines are helpless on hatching and are entirely dependent on their parents for food and shelter (altricial) for several weeks.
As the birds evolved and diversified, they tended to become smaller and also more altricial. This reduction in body size and increased altriciality are reflected in the genomes. It was these transitional changed that the researchers detected.
These major changes were occurring within 3-5 million years after the mass extinction.
In case any creationists are still under the delusion that mainstream biologists are abandoning the scientific Theory of Evolution (TOE) in favour of their childish magical story involving a magic man made of nothing who magicked everything into existence out of nothing with some magic words, here is an example of how the TOE is used to understand and make sense of the observable facts.
It addresses the question of how the ability to learn and imitate sounds evolved in birds.
Basically, ornithologists had thought that birds could be divided into two groups - those which can learn sounds (parrots, songbirds, and hummingbirds) and those which can’t - and that this ability in the former group had evolved sometime after modern birds had diversified from their avian dinosaur ancestors, but the fact that a small New Zealand bird, the titipounamu or rifleman, Acanthisitta chloris, has the rudiments of this ability suggests it may have been present in the common ancestor of both groups.
In other words, the ability to learn and imitate sounds may be evidence of common descent.
Not only is there no sign at all that the conversion of leading biologists to creationism's childish fairy tale in place of the scientific Theory of Evolution, that creationist cult leaders have been assuring their dupes is imminent - and has been for the last 50 years or so - it's as though they've never even heard of it. They still keep finding evidence of events that happened ten of thousand, even tens of millions of years before the Universe existed, according to the ludicrous creation myths they are supposedly about to adopt. And those events are frequently evidence of evolution or recent human history.
Imagine a serious, grown-up scientist who has been through university, with so much contrary information at his or her fingertips believing that account of magic creation 10,000 years ago written by ignorant Bronze Age pastoralists yet belonging to the creation cult requires fools to believe such an absurdity.
About 9 months ago while on holidat near Bezier, France, we were driving to Carcassonnes when, about a mile apart, we saw two majestic eagles that neither of us recognised. I now know they were Bonelli's eagles, and I probably have the activities of early Homo sapiens about 40,000 years ago to thank for them being there.
A study led by scientists from the University of Granada (UGR), Spain, shows how the activities of early Homo sapiens in the Iberian Peninsula, 40,000 years before creationists think their god created a universe consisting of a small, flat planet with a dome over it in the Middle East, may well have facilitated the expansion of the range of Bonelli's Eagle, Aquila fasciata, north of the Mediterranean Basin.
This paper is enough to send a dedicated creationist disinformation specialist into intellectual summersaults. On the one hand, they hate the fact that dinosaurs lived so long ago and are evidence of life on earth tens and hundreds of millions of years ago, so they bend over backwards to prove they were around just a few thousand years ago, and then, horror of horrors, along come some clever scientists and show evidence that some dinosaurs even evolved into birds!
And now, courtesy of Palaeontologists at University College Cork (UCC) in Ireland , we have evidence of the transition from scales to feathers in pre-avian dinosaurs.
But it gets even worse! Creationists frauds have ben claiming for several years now that 'fossilised' soft dinosaur tissues prove they were just a few thousand years old because, despite the perfectly rational explanation of how 'soft' tissues can be preserved for many millions of years in the right circumstances, like the presence of iron, the evidence of this transition from scales to feathers would found in fossilised soft tissue!
So, a creationist disinformation specialist must now argue that fossilised dinosaur soft tissues don't exist, and when they do, they prove dinosaurs lived recently. One problem they face is that the fossilised skin isn't skin at all, or even the fibrous protein parts of skin; it has been entirely mineralised aa silicate, preserving only the morphology, including the surface patterning - which is where the evidence lies.
The University College Cork scientists have just published their findings, open access, in Nature Communications and in a UCC news release:
Ducks, like almost all taxonomic categories, are a nightmare for creationists because they aren't mentioned in the Bible so it's not possible to put them into their primitive 'kind' classification, other than saying they are some 'kind' of bird, or maybe a 'duck kind'. The problem is there are so many different families of duck and several different species within each family. Some of them look very different to other ducks and have clearly evolved unique characteristics such as the long, serrated beak of the fish-eating mergansers, which places them in a different clade to the dabbling diving and torrent ducks.
By any definition then, evolving from a basic 'duck' into a merganser is the evolution of a new taxon, or 'macro-evolution' as creationists like to call it, while proclaiming it to be impossible because of some undefined magical process that intervenes to prevent it.
Which leaves creationists with nowhere to go but to claim mergansers were on the Ark, since they couldn't have evolved from a parent pair of 'ducks' at the abnormal warp-speed rate of evolution, that no-one noticed or thought to record, that creationists now claim must have happened in the few years since their favourite genocidal flood, that left no trace of itself.
But that's nothing compared to the genetic evidence that shows how, for example, the mergansers, evolved and spread geographically 18 million years before creationists believe there was a universe and again between 14 and 7 million years before creationists' god decided to create a small flat planet with a dome over it in the Middle East, 10,000 years ago, that creationists still believe accurately describes all that there is.
First, a little AI information about mergansers:
Tell me all about mergansers, their distribution and evolutionary origins, please. Mergansers are a group of ducks belonging to the genus Mergus, which is part of the family Anatidae. They are known for their distinctive serrated bills, which are well-adapted for catching fish, their primary diet. Here’s a detailed overview of mergansers, including their distribution, characteristics, and evolutionary background.
Characteristics
Mergansers are medium to large-sized ducks with slender bodies and long, pointed bills. Their bills are equipped with saw-like edges that help them grasp slippery fish. There are several species within the Mergus genus, including:
Common Merganser (Mergus merganser): Also known as the goosander, this species has a widespread distribution across North America, Europe, and Asia.
Red-breasted Merganser (Mergus serrator): Found in North America, Europe, and Asia, this species prefers coastal and large inland waters.
Hooded Merganser (Lophodytes cucullatus): Native to North America, it is smaller than other mergansers and has a distinctive fan-shaped crest.
Brazilian Merganser (Mergus octosetaceus): One of the rarest mergansers, it is found in central Brazil and is critically endangered.
Scaly-sided Merganser (Mergus squamatus): This species inhabits parts of East Asia and is also considered endangered.
Distribution
Mergansers have a wide distribution, with different species occupying various regions around the globe:
Common Merganser: Breeds in forested regions near lakes and rivers across northern North America, Europe, and Asia. In winter, they migrate to more temperate areas.
Red-breasted Merganser: Found in boreal and temperate regions across the Northern Hemisphere. They often winter in coastal waters, estuaries, and large inland lakes.
Hooded Merganser: Prefers forested wetlands, rivers, and ponds in North America. They are non-migratory in some parts of their range but move to ice-free waters in winter.
Brazilian Merganser: Restricted to central Brazil, particularly in the Serra da Canastra region. Their habitat includes clear, fast-flowing rivers.
Scaly-sided Merganser: Breeds in forested rivers of northeast China and the Russian Far East. They migrate to southern China, Korea, and Japan for winter.
Evolution
Mergansers belong to the Anatidae family, which includes ducks, geese, and swans. The evolution of mergansers is part of the broader adaptive radiation of the Anatidae, which diversified to exploit a variety of ecological niches. The serrated bills of mergansers are a specialized adaptation for piscivory (fish-eating), differentiating them from other ducks.
Evolutionary Background
Ancestral Origins: The Anatidae family likely originated in the late Cretaceous to early Paleogene periods. Early diversification allowed these birds to adapt to various ecological roles, including herbivory, omnivory, and piscivory.
Adaptive Radiation: The diversification of Mergus species is an example of adaptive radiation, where different species evolved to occupy different ecological niches, particularly in terms of habitat preference and diet specialization.
Morphological Adaptations: The distinctive serrated bills of mergansers are a key evolutionary trait that sets them apart from other ducks. This adaptation is crucial for their fish-based diet.
Conservation Status
Several species of mergansers are of conservation concern:
Brazilian Merganser: Critically endangered, with an estimated population of fewer than 250 individuals in the wild. Habitat destruction and water pollution are major threats.
Scaly-sided Merganser: Endangered, with threats including habitat loss due to logging and river damming, as well as pollution and human disturbance.
Conclusion
Mergansers are a fascinating group of ducks with specialized adaptations that enable them to thrive as piscivores in a variety of aquatic habitats. Their wide distribution and evolutionary history reflect the dynamic processes of diversification and adaptation in the Anatidae family. Conservation efforts are crucial for the survival of the endangered species within this group, highlighting the need for habitat protection and restoration.
The evidence of when the now extinct New Zealand merganser got to New Zealand and how it is related to other mergansers is the subject of an article in The Conversation by Nic Rawlence, Senior Lecturer in Ancient DNA, and Alexander Verry, Researcher, both of the University of Otago, New Zealand. Their article is reprinted here under a Creative Commons license, reformatted for stylistic consistency:
Ancient DNA from an extinct native duck reveals how far birds flew to make New Zealand home
Auckland Island merganser. Artistic reconstruction by J. G. Keulemans from Bullers Birds of New Zealand (1888)
Ask a bird lover if they have heard of the extinct giant moa or its ancient predator, Haast’s eagle, and the answer will likely be yes. The same can’t be said of New Zealand’s extinct, but equally unique, mergansers – a group of fish-eating ducks with a serrated bill.
The only southern hemisphere representatives of this group are the critically endangered Brazilian merganser and those from the New Zealand region, which are now extinct.
Unlike some of New Zealand’s other extinct birds, the biological heritage of our enigmatic mergansers is shrouded in mystery. But our new research on the extinct Auckland Island merganser is changing the way we think about the origins of New Zealand’s birds. Did the ancestors of the merganser come from South America or the northern hemisphere – and when did they arrive?
Mergansers are relatively common in the northern hemisphere but were limited to Brazil and the New Zealand region in the southern hemisphere.
Author provided
Lost to humans and pests
Mergansers were spread across the three main islands of New Zealand at the time of Polynesian arrival in the 13th century, as well as the Auckland Islands to the south and the Chatham Islands to the east.
Over-hunting, habitat destruction, and predation from the Pacific rat and Polynesian dog resulted in the extinction of mergansers on the New Zealand mainland and the Chatham Islands. By the time Europeans arrived in the 17th century, mergansers were restricted to an isolated population on the sub-Antarctic Auckland Islands.
European discovery of the Auckland Islands in 1806 led to a formal description of the Auckland Island merganser in 1841. However, European discovery brought new predators like pigs and cats.
Mergansers were also sought after as specimens for the museum trade. The last known Auckland Island merganser was shot and collected in 1902, a mere 61 years after its discovery.
Auckland Island Merganser, Mergus australis, collected June 1902, Auckland Islands, New Zealand.
Only recently have merganser bones from the Chatham Islands been described as a distinct species. The distinction of the Chatham Island merganser hints at greater merganser diversity in the New Zealand region than previously thought. It is possible the merganser bones found on mainland New Zealand may be another species, but only ongoing research will be able to confirm this.
From the northern hemisphere to NZ
The extinction of mergansers from the New Zealand region has meant their evolutionary history has remained a mystery. Did their ancestors, and those of the Brazilian merganser, arrive via independent colonisation events from the northern hemisphere? Or was there a single push into the southern hemisphere, followed by subsequent divergence events?
To find out more, we sequenced ancient DNA from an Auckland Islands merganser and a Brazilian merganser. This allowed us to reconstruct the evolutionary history of the wider group.
We found mergansers originated in the northern hemisphere, diverging from their closest relatives some 18 million years ago, before rapidly evolving into several different species between 14 and seven million years ago.
The mergansers from the New Zealand region are most closely related to the northern hemisphere common merganser. Their ancestors arrived here at least seven million years ago in a separate colonisation event to the one that gave rise to the Brazilian merganser.
Further genetic research is currently underway. The goal is to reconstruct the evolutionary history of mergansers within the New Zealand region.
The global origins of New Zealand’s birds
Many New Zealanders believe the country’s native birds originate from Australia. Increasingly though, genetic and palaeontological research shows a number of our feathered friends hail from further afield.
The long journey of blue-eyed shags started in South America, with the birds island hopping via Antarctica and the sub-Antarctic islands to New Zealand. Mergansers arriving from the northern hemisphere add another piece to the puzzle.
It is possible that fossils of extinct mergansers (and other birds with distant geographic origins) will be discovered as palaeontologists increasingly focus on previously neglected and newly discovered southern hemisphere fossil deposits.
Only then, combined with the power of ancient DNA, will we be able to fully understand how New Zealand’s dynamic geological, climatic and human history has influenced the colonisation and diversification of birds on this isolated South Pacific archipelago. Nic Rawlence, Senior Lecturer in Ancient DNA, University of Otago and Alexander Verry, Researcher, Department of Zoology, University of Otago
Abstract
Mergansers are riverine and coastal piscivorous ducks that are widespread throughout North America and Eurasia but uncommon in the Southern Hemisphere. One species occurs in South America and at least two extinct species are known from New Zealand. It has been proposed that these Southern Hemisphere merganser lineages were founded by at least two independent dispersal events from the Northern Hemisphere. However, some morphological and behavioural evidence suggests that Southern Hemisphere mergansers may form a monophyletic clade that descended from only a single dispersal event from the Northern Hemisphere. For example, Southern Hemisphere mergansers share several characteristics that differ from Northern Hemisphere mergansers (e.g. non-migratory vs. migratory, sexual monochromatism vs. sexual dichromatism, long vs. short pair bonds). We sequenced complete mitogenomes from the Brazilian merganser and an extinct merganser from New Zealand—the Auckland Island merganser. Our results show that the Brazilian and Auckland Island mergansers are not sister-taxa, and probably descend from two separate colonization events from the Northern Hemisphere at least 7 Mya. Nuclear (palaeo)genomic data may help to further resolve the relationship between living and extinct mergansers, including merganser fossils from New Zealand that have not been subjected to palaeogenetic analysis.
INTRODUCTION
Mergansers (Mergus spp.) are a group of riverine and seasonally coastal fish-eating ducks that have a widespread Northern Hemisphere distribution but are uncommon in the Southern Hemisphere (Kear 2005, Williams et al. 2012, 2014) (Fig. 1). They are characterized by a serrated bill, and include the endangered scaly-sided merganser (M. squamatus Gould 1864) from north-east Asia; the common merganser (M. merganser Linnaeus 1758), and the red-breasted merganser (M. serrator Linnaeus 1758), which have widespread Northern Hemisphere distributions; the critically endangered Brazilian merganser (M. octosetaceus Vieillot 1817); and two currently recognized extinct species from the New Zealand region—M. australis Hombron and Jacquinot 1841 and M. milleneri Williams and Tennyson 2014 from the Auckland and Chatham Islands, respectively. While the hooded merganser Lophodytes cucullatus (Linnaeus 1758), previously M. cucullatus, from North America has a serrated bill, it is not considered a ‘true’ merganser (e.g. Buckner et al. 2018, Lavretsky et al. 2021). The taxonomic relationship of the smew Mergellus albellus (Linnaeus 1758) from Eurasia is currently unresolved; it is sometimes suggested to be more closely related to Mergus and Lophodytes or to goldeneyes (Bucephala spp.) (Livezey 1995, Buckner et al. 2018, Lavretsky et al. 2021).
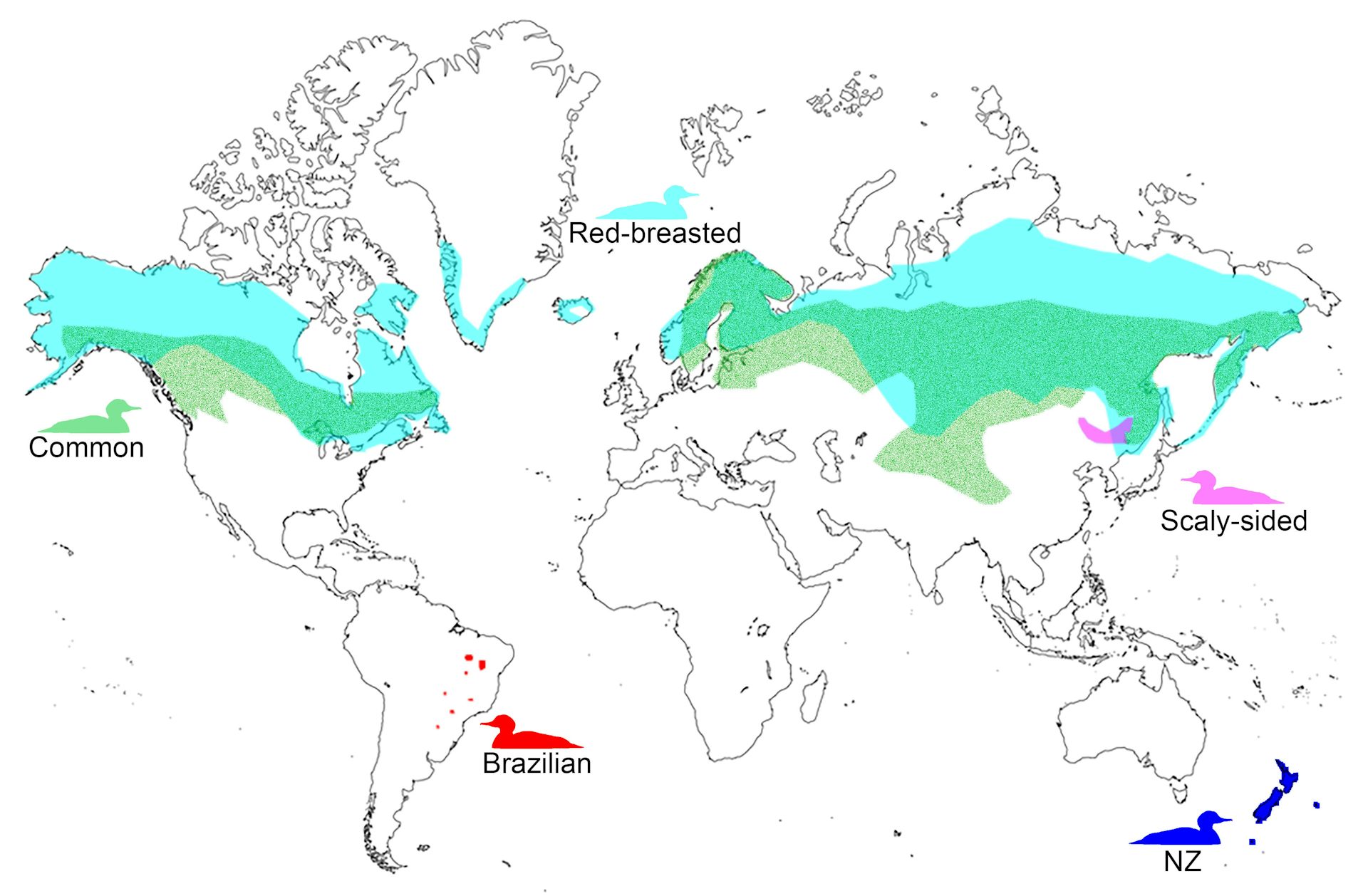
Figure 1.
Schematic of the breeding distributions of Mergus spp. The New Zealand (NZ) lineage encompasses the Auckland Island merganser (465 km south of NZ) and Chatham Island merganser (785 km east of NZ), as well as Mergus spp. from mainland NZ. Breeding distributions are based off the Cornell Lab of Ornithology Birds of the World website.
The now extinct Auckland Island merganser M. australis (or miuweka) (Fig. 2) was formally described in 1841, based on a specimen collected on the sub-Antarctic Auckland Islands, 465 km south of mainland New Zealand. Rare Late Holocene-aged merganser bones have been found in coastal sand dune deposits (including Māori middens) on New Zealand’s three main islands (Stewart, North, and South), and the Auckland and Chatham Islands (Tennyson and Martinson 2007, Williams et al. 2014, Tennyson 2020). Bones from the latter were recently described as a distinct species M. milleneri, which was smaller than the nominate M. australis, with a shorter skull, relatively shorter premaxilla, smaller sternum and keel, relatively shorter wing bones, and a narrower pelvis (Williams et al. 2014). The taxonomic status of merganser bones from mainland New Zealand is unresolved (i.e. cannot be assigned to either M. australis or M. milleneri), and are currently recognized as Mergus spp. (Birds New Zealand Checklist Committee 2022).
Figure 2.
In the Southern Hemisphere, mergansers are only known from the New Zealand region and South America, represented here by the Auckland Island merganser. A, artistic reconstruction by J.G. Keulemans from Buller (1888); B, historical museum skin (Museum of New Zealand Te Papa Tongarewa OR.001357); C, the Brazilian merganser (photo by Savio Freire Bruno CC BY-SA 3.0).
Mergansers in the New Zealand region are mainly thought to have occupied a riverine and seasonally coastal habitat (e.g. sheltered bays during winter; Kear 2005, Williams et al. 2012, 2014). It is likely that they mainly nested in tree cavities, but also caves in some instances, as the remains of adults, chicks, and eggs have been found within a cave on the Chatham Islands (Williams et al. 2014). By the 17th century, mergansers had been extirpated from the Chatham Islands and mainland New Zealand, and survived only on the Auckland Islands. A combination of subsistence hunting, and predation from the Pacific rat (Rattus exulans) and Polynesian dog (Canis familiaris), probably led to the extinction of mergansers across most of their prehistoric range (Tennyson and Martinson 2007, Greig and Rawlence 2021.1). On the Auckland Islands, predation from introduced pigs (Sus scrofa) and cats (Felis catus), and collecting for the museum trade, resulted in their extinction—indeed the last known Auckland Island merganser specimen was shot and collected in January 1902 (Williams 2012.1).
The only extant merganser in the Southern Hemisphere—the critically endangered Brazilian merganser (Fig. 2)—is one of the rarest birds in the world, comprising only 250 wild individuals. It is split across three remnant populations in Brazil, but once had a more widespread historical distribution encompassing Argentina and Paraguay (Vilaca et al. 2012.2, Maia et al. 2020.1). The Brazilian merganser has undergone a significant population bottleneck, yet different remnant populations can still be genetically identified (Maia et al. 2020.1). Like mergansers from the New Zealand region, the Brazilian merganser occupies riverine habitats, and often nests in tree cavities or rock crevasses (Vilaca et al. 2012.2, Maia et al. 2020.1).
It has been proposed that the Southern Hemisphere mergansers were founded by independent dispersal events to the New Zealand region and South America from the Northern Hemisphere (e.g. Livezey 1995). Based on behavioural characteristics, Johnsgard (1961) tentatively assigned the Brazilian merganser as sister-species to a clade comprising the other Mergus species, with the Auckland Island merganser as the sister-species of the common merganser and scaly-sided merganser. In contrast, using morphological characters, Livezey (1989, 1995) assigned the Auckland Island merganser, then Brazilian merganser, as successive sister-species to all other Mergus species, though with weak to moderate bootstrap support. Using mitochondrial DNA (mtDNA) sequences, Buckner et al. (2018) suggested the Brazilian merganser was the sister-species to the scaly-sided merganser, albeit with weak support. However, some evidence suggests that Southern Hemisphere mergansers may be closely related to one another, potentially even sister-species, as they share several behavioural (e.g. non-migratory and long pair bonds) and morphological (e.g. sexually monochromatic) characteristics, in contrast to their Northern Hemisphere congeners (e.g. migratory, short pair bonds, and sexual dichromatism; Livezey 1995). In addition, recent genetic studies of other extinct Southern Hemisphere avian species have also revealed unexpected evolutionary connections between birds from New Zealand, South America, and Africa (e.g. Mitchell et al. 2014.1a, 2014.2b, Boast et al. 2019, Rawlence et al. 2022.1, Verry et al. 2022.2a). As such, the phylogenetic relationships of the Southern Hemisphere mergansers, when their ancestors arrived in the region, and from where, remain unresolved.
In this study, the first genetic study of a New Zealand Mergus species, we sequenced mitochondrial genomes (mitogenomes) from historical museum specimens from the Auckland Island merganser and Brazilian merganser, and analysed them within a phylogenetic framework of Mergini mitogenomes (Liu et al. 2012.3, Lavretsky et al. 2021). These data were used to determine the phylogenetic relationships and divergence dates within mergansers.
Creationists needn't be too embarrassed by these facts about the distribution and evolution of mergansers, after all, the simple Bronze Age Canaanites who made up the origin myths to fill the gaps in their knowledge and understanding, could have known nothing of them, since they are largely absent from the Middle East, nor of southern hemisphere land masses such as South America, Australia and New Zealand when they didn't even realise there is a southern hemisphere, or of the ducks and their families.
And, of course, they knew nothing about the history of Earth and the life on it, so how could they be expected to have known about the evolutionary history and geographic distribution of these ducks? With their long, serrated beaks and fish diet, they probably wouldn't have recognised mergansers as a type of duck even.
All they had to draw on was their limited knowledge of the small scrap of the globe they inhabited, which is why they got almost everything they described so hopelessly wrong. The lesson from Genesis is just how badly you do when you know little and depend on guesswork from the point of view of someone with the knowledge understanding of a pre-school toddler of today.