

Homo floresiensis and dwarf elephants went extinct at about the same time.
Image: GETTY.
Science is a continuous process of reconciling new discoveries and existing theories, and revising those theories as necessary. It's a process that often takes time, sometimes a long time, involving reassessing the new discovery, formulating new hypotheses and then waiting for more discoveries to provide enough evidence to falsify some hypothese and add weight to remaining ones, until a consensus is reached that one hypothesis has emerged the winner by failing to be falsified and as having the best fit with the known facts.
Creationists, who base their beliefs not on evidence, but on dogma and 'faith', i.e., believing what someone told them to believe without any evidence, find this process baffling because it looks like scientists can't agree and keep changing their minds. But one thing scientists don't do, unlike theologians, especially in the past before basic human decency got incorporated into religion, is kill one another and have campaigns of persecution and ostracism, while dividing into warring factions with neither side havign any evidence with which to resolve disputes.
What causes miniaturisation in an island population, ege, Homo floresiensis? Miniaturization in island populations, also known as the "island rule" or "Foster's rule," is a phenomenon where species that are isolated on islands often exhibit a significant change in size compared to their mainland counterparts. In the case of Homo floresiensis, also known as the "Hobbit," this phenomenon led to a much smaller stature compared to other hominins. Several factors contribute to this miniaturization:If scientists behaved that way we would now have tens of thousands of different groups of mutually hostile scientists each with a different account of human origins and each appointing its professors and doctors but not recognising the others, just as there are tens of thousands of different, mutually hostile protestant churches in the USA, each believing they are the one true church and all the others are heretical.
- Limited Resources: Islands typically have limited food and other resources. Smaller body sizes require fewer resources, making it advantageous for survival in such environments. This is particularly critical during periods of food scarcity.
- Predation Pressure: The absence or reduction of predators on islands can also lead to size reduction. Without the need to be large for defense, smaller sizes can evolve as they are more efficient in resource usage.
- Ecological Niches: On islands, species often need to adapt to different ecological niches than their mainland relatives. This can lead to changes in size as species adapt to new diets, behaviors, and lifestyles.
- Genetic Drift and Founder Effect: Island populations are often small and isolated, leading to a limited gene pool. Genetic drift and the founder effect (when a new population is established by a very small number of individuals) can cause rapid changes in traits, including body size.
- Developmental and Physiological Constraints: In some cases, smaller body sizes may result from developmental and physiological constraints that favor reduced growth rates under the environmental conditions prevalent on islands.
- Thermoregulation: Smaller bodies are more efficient in certain climates, especially in warmer climates where smaller size can help with heat dissipation.
Homo floresiensis is a prime example of island dwarfism. This species lived on the island of Flores in Indonesia and exhibited a significantly smaller stature than contemporary Homo sapiens. Fossil evidence suggests that adult H. floresiensis individuals stood about 1.1 meters (3.6 feet) tall. The combination of limited resources, absence of large predators, and other ecological factors likely contributed to this pronounced miniaturization.
Science, by contrast, is entirely evidence-led, so evidence is the court of first, last and final resort. Whatever the evidence shows, is what science accepts.

Artist's impression of H. floriensis compared to other hominins.
From left to right: Homo habilis, Homo Sapiens, Homo floresiensis, Homo Erectus, Paranthropus boisei, Homo heidelbergensis and Homo neanderthalensis.
From left to right: Homo habilis, Homo Sapiens, Homo floresiensis, Homo Erectus, Paranthropus boisei, Homo heidelbergensis and Homo neanderthalensis.
Back in 2003 (how time flies!) a team of palaeontologists co-led by the late Mike Morwood, discovered fossils of a previously unknown hominin in the Liang Bua cave on the Indonesian island of Flores. It was unlike any other homin, in that is appeared to be an adult but about the height of a child and a small brain, a dwarf hominin in Indonesia! They named it Homo floresiensis but the populr press soon named it 'The Hobbit' after J.R.R.Tolkein's book of thr same name.
Minaturisation is frequently found on islands, and indeed Flores had its own population of dwarf elephants too, but the questions were, firstly, was it really a normal healthy adult, and secondly what did it descend from?
The various hypotheses were:
- It was a modern H. sapiens with a pathological condition.
- It was a dwarfed descendant of H. erectus from nearby Java
- It had evolved from a more primitive African hominin such as H. habilis, or even and Australopithecine, such as Au. afarensis (Lucy) which had migrated out of African in an even earlier migration that H. erectus and H. sapiens.
The discovery, made in 2013, and its significance to the debate is the subject of an open access paper in Nature Communications and is explained by three of the archaeologists in an article in The Conversation. They are Adam Brumm, Professor of Archaeology, Griffith University, Australia, Gerrit (Gert) van den Bergh, Researcher in palaeontology, University of Wollongong, Australia and Yosuke Kaifu, Professor of Biological Anthropology, The University Museum, University of Tokyo, Japan. Their article is reprinted here under a Creative Commons license, reformatted for stylistic consistency:
Tiny fossil arm bone sheds light on evolution of ancient Indonesian ‘hobbits’
The arm bone fragment excavated in 2013 at the site Mata Menge.
Adam Brumm, Griffith University; Gerrit (Gert) van den Bergh, University of Wollongong, and Yosuke Kaifu, University of TokyoY. Kaifu
We have discovered rare and very old human fossils on the Indonesian island of Flores, including an astonishingly small adult limb bone. Our finds are reported in a new paper out today in Nature Communications.
Dated to around 700,000 years old, these findings throw new light on the evolutionary history of Homo floresiensis, the so-called “hobbits” of Flores who inhabited this small island east of Bali as recently as 50,000 years ago.
A strange new human
In 2003, an archaeological dig co-led by the late Australian–New Zealand archaeologist Mike Morwood unearthed the fossils of a previously unknown species of early human at Liang Bua cave on Flores. Named Homo floresiensis, these humans were extremely short, with tiny brains and a host of unusual features.
Nothing like this had ever been found before, so the origins of the creature were disputed. Scientists argued over whether the remains belonged to a new human species or a modern person with a pathological condition. One professor even claimed the “hobbit” had a recent dental filling.
More sober-minded assessments suggested Homo floresiensis was a dwarfed descendent of Asian Homo erectus from nearby Java. Other palaeoanthropologists argued Homo floresiensis had evolved from an earlier and more primitive African hominin, perhaps Homo habilis or Australopithecus afarensis (or “Lucy”).
Digging for pre-“hobbit” ancestors
In an attempt to resolve this puzzle, we searched for fossils of “hobbit” ancestors in the So’a Basin. This is an area of tropical grasslands east of Liang Bua where stone artefacts dating to at least a million years ago have been found.
In 2016, after years of digging at a site named Mata Menge, we reported the first early human fossils from outside Liang Bua: several isolated teeth and a jaw fragment found in a layer of sandstone around 700,000 years old.

The So'a Basin of central Flores where the oldest known hominin fossils from the island were found in a sandstone layer at Mata Menge dating to 700,000 years old. A nearby site has yielded stone tools dating to at least one million years ago.
A. Brumm

The dig at Mata Menge, where fossils of extinct animals occur alongside stone artefacts in hard sedimentary layers deposited by small streams. The fossil deposits date to between around 700,000 years ago to as early as 773,000 years ago. In the foreground are tusks and limb bones of stegodonts, an extinct relative of elephants.
A. Brumm
It was also unclear what species the fossils belonged to, owing to the limited skeletal elements at hand. However, some of the Mata Menge teeth seemed to be intermediate in form between early Asian Homo erectus and Homo floresiensis.
The new fossils
In 2013, before the teeth and jaw fragment, we had found a tiny limb bone fragment broken into several pieces in the 700,000-year-old fossil layer at Mata Menge.
We initially thought it was from a reptile, until in 2015 the curator of fossils at the Geology Museum in Bandung, Indra Sutisna, restored the bone fragment and recognised it as the shaft of a human upper arm bone. This was later confirmed by one of us (Kaifu), although initially it was thought to belong to a child because of its extremely small size.

The 700,000-year old adult humerus fragment from Mata Menge.
Y. Kaifu
The fossil limb bone provides a wealth of evidence about the body size evolution of Homo floresiensis.
The microscopic structure of this tiny bone indicates it came from an adult. Based on the estimated length of the bone (206–226mm), we were able to calculate this hominin was about 100cm tall.
A skeleton of 60,000-year-old Homo floresiensis, the type specimen found at Liang Bua, was estimated to be around 106cm tall, based on the length of its thigh bone. (Based on its arm bones, its height would be estimated at around 111.5cm.)
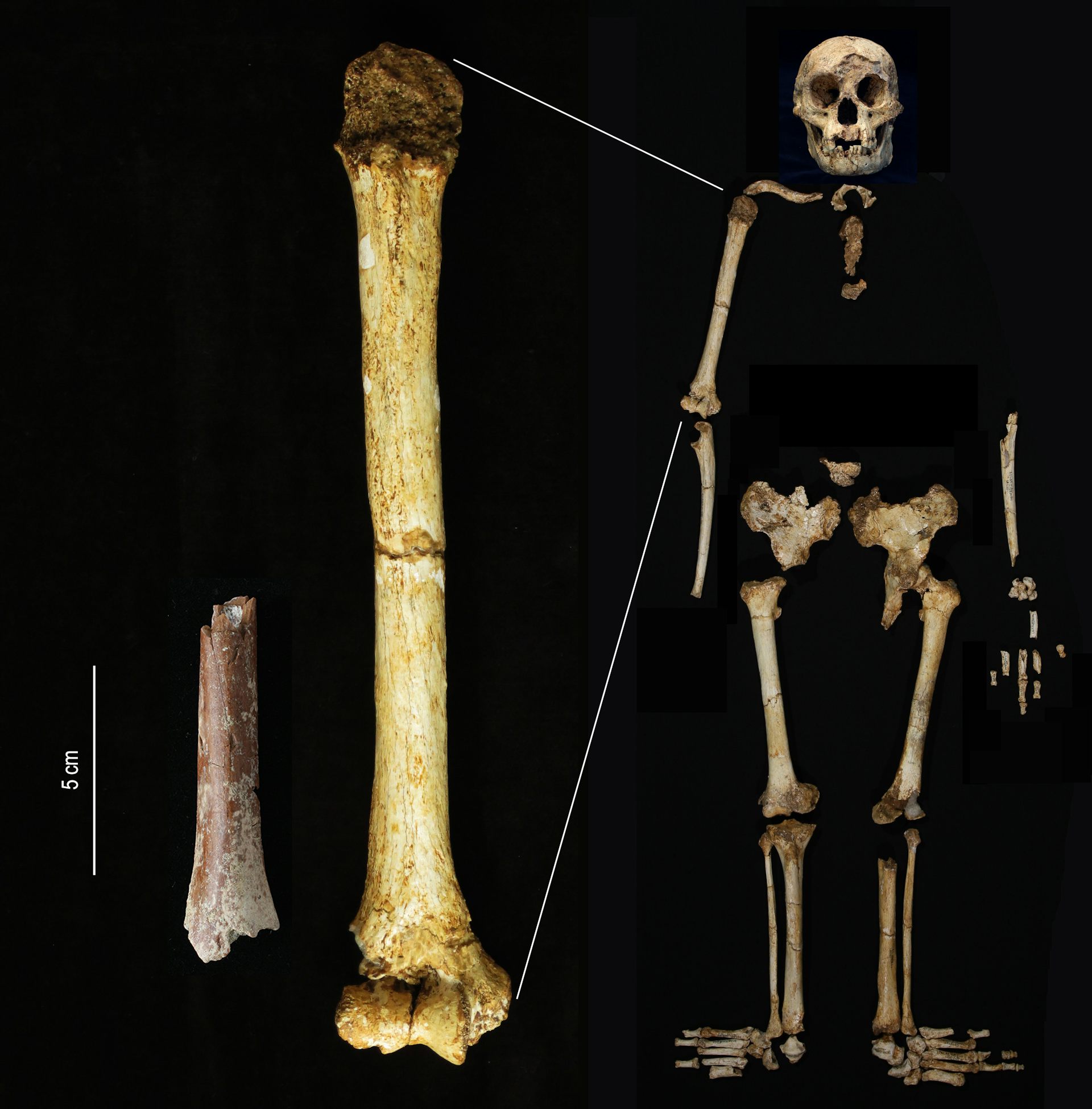
The Mata Menge humerus fragment (left) shown at the same scale as the humerus of Homo floresiensis from Liang Bua.
Y. Kaifu
Two additional hominin teeth from Mata Menge are also small in size and one bears shape characteristics most consistent with early Homo erectus of Java. This does not support the hypothesis that Homo floresiensis evolved from a pre-erectus hominin, the likes of which have never been found in Southeast Asia.
The beginnings of Homo floresiensis
The Mata Menge human remains, which now total ten fossil specimens, are from at least four individuals (including two children). The teeth are rather similar anatomically to those of the Liang Bua Homo floresiensis, as is the arm bone fragment. Hence the Mata Menge hominin should probably be thought of as an older variant of this hominin – though its teeth do not have some of the changes found in the latter Liang Bua Homo floresiensis.
It is certainly evident that extreme body size reduction occurred very early in the history of the Flores hominins – by at least 700,000 years ago.
We should note that the Mata Menge arm bone is not necessarily Homo erectus-like. It more resembles small-bodied Homo such as Homo floresiensis and Homo naledi.
But the Liang Bua hominins were very odd quirks of evolution. They display a mixture of archaic and modern traits, some that hint at their descent from Homo erectus and others that imply they evolved unique characters over long isolation.
The latter include not only small body and brain sizes but also apparently primitive limb proportions (a combination of longer arms and shorter legs) as well as advanced or “hyper-modern” molar morphology.
In our view, the new fossils from Mata Menge confirm the hypothesis that a group of early Asian Homo erectus somehow became isolated on Flores and underwent a remarkable process of evolutionary change, giving rise to Homo floresiensis.
But the fossils are few and fragmentary and hence the history of the Flores hominins is not known with certainty. The picture may change dramatically if we are lucky enough to find a partial skeleton or even a more substantial portion of a skull at Mata Menge or elsewhere in the So'a Basin.
Adam Brumm, Professor of Archaeology, Griffith University; Gerrit (Gert) van den Bergh, Researcher in palaeontology, University of Wollongong, and Yosuke Kaifu, Professor of Biological Anthropology, The University Museum, The University of Tokyo, University of Tokyo
This article is republished from The Conversation under a Creative Commons license. Read the original article.

AbstractOf course, this evidence is entirely inconsistent with the Bible narrative, both in it timescale and in it daft notion of a single hominin ancestor 10,000 years ago, magicked into existence without ancestors. Early, archaic hominins had dispersed out of Africa and diversified into a number of regional varieties, sub-species and species, long before anatomically-modern humans migrated into Eurasia, where they met and interbred with the descendants of those earlier migrants.
Recent discoveries of Homo floresiensis and H. luzonensis raise questions regarding how extreme body size reduction occurred in some extinct Homo species in insular environments. Previous investigations at Mata Menge, Flores Island, Indonesia, suggested that the early Middle Pleistocene ancestors of H. floresiensis had even smaller jaws and teeth. Here, we report additional hominin fossils from the same deposits at Mata Menge. An adult humerus is estimated to be 9 − 16% shorter and thinner than the type specimen of H. floresiensis dated to ~60,000 years ago, and is smaller than any other Plio-Pleistocene adult hominin humeri hitherto reported. The newly recovered teeth are both exceptionally small; one of them bears closer morphological similarities to early Javanese H. erectus. The H. floresiensis lineage most likely evolved from early Asian H. erectus and was a long-lasting lineage on Flores with markedly diminutive body size since at least ~700,000 years ago.
Introduction
The So’a Basin in central Flores, Indonesia, is a key region for elucidating the origin and evolution of H. floresiensis, a diminutive hominin species known from the Late Pleistocene at Liang Bua, a limestone cave in western Flores1,2. As with another small-bodied Homo discovered in Luzon3, the evolutionary history of this insular hominin species has been the subject of protracted debate4. Previous field studies of the Early to Middle Pleistocene (Calabrian–Chibanian) sequence of the So’a Basin have recovered fossil remains of endemic fauna (dwarfed Stegodon, Komodo monitor, giant rat, birds, crocodiles and tortoises)5,6, technologically simple stone artefacts (the oldest of which date back to at least 1.02 ± 0.02 million years ago [Ma])7,8, and, importantly, a fragmentary mandible and six isolated teeth of a small-sized hominin9. These hominin fossils were excavated from a sandstone layer of fluvial origin (Layer II) of the upper fossil-bearing interval at the Mata Menge site, which is dated to between 0.65 and 0.773 Ma5,6. These fossils exhibit general morphological affinities to the type series of H. floresiensis from Liang Bua (0.1–0.06 Ma)10 and to early H. erectus from Java (1.1–0.8 Ma)11, but lack the unique molar specializations characterizing the former and were substantially smaller than the latter9.
Overall, the Mata Menge fossils suggest that they represent an ancestral segment of the Liang Bua H. floresiensis lineage, and that the Flores hominins are dwarfed descendants of large-bodied early Asian H. erectus9. Some cladistic/phylogenetic analyzes, however, support a direct evolutionary link between H. floresiensis and smaller-bodied basal Homo such as H. habilis or even Australopithecus12,13,14. It is important to resolve this controversy in order to elucidate the pattern and timing of body size evolution in the Flores hominins.
Notably, the Mata Menge mandible and teeth are slightly smaller than the type specimens of H. floresiensis from Liang Bua. This suggests that drastic dentognathic reduction had occurred on Flores by the early Middle Pleistocene epoch, more than 600,000 years before the earliest fossil evidence for H. floresiensis at Liang Bua. Until now, however, the lack of postcranial elements in the Mata Menge assemblage had limited our understanding of body size evolution on Flores.
In this paper, we report the discovery and morphology of a hominin postcranial fossil from Mata Menge, an extremely small distal humeral shaft (SOA-MM9) (Fig. 1). This specimen and two small-sized teeth (SOA-MM10 and SOA-MM11) were recovered as additions to the existing hominin assemblage from Layer II at this site (Table 1). Our histomorphic examination confirms the adult status of the humerus. We also show that shaft morphology is more similar to small-bodied Homo (e.g., LB1 and H. naledi) than to Australopithecus (e.g., A.L. 288-1), and a molar crown (SOA-MM11) bears closer shape similarities to early Javanese H. erectus than to early African Homo. The increased Mata Menge fossil sample supports its classification to an early representative of H. floresiensis, which probably experienced drastic body size reduction from large-bodied Asian H. erectus sometime between ~1.0 and 0.7 Ma.ResultsFig. 1: New fossils from Mata Menge. a–f SOA-MM9 humerus in anterior, lateral, posterior, medial, proximal, and distal views. g Micro-CT section of SOA-MM9 at the distal end indicated in (c). h and i LB1 humerus in anterior and medial views. Note the hollowed area on the posterior surface of the distal end (filled triangles in f and g), which serves as an osteometric landmark (hOF point). Abbreviations: ant = anterior, post = posterior, lat = lateral, med = medial, DT = deltoid tuberocity, NF = nutrient foramen, HS = location for histological section. j SOA-MM10 right dc. From left to right, occlusal, labial, and lingual views. k SOA-MM11 left M3. From left to right, occlusal, buccal, and lingual views.
a–f SOA-MM9 humerus in anterior, lateral, posterior, medial, proximal, and distal views. g Micro-CT section of SOA-MM9 at the distal end indicated in (c). h and i LB1 humerus in anterior and medial views. Note the hollowed area on the posterior surface of the distal end (filled triangles in f and g), which serves as an osteometric landmark (hOF point). Abbreviations: ant = anterior, post = posterior, lat = lateral, med = medial, DT = deltoid tuberocity, NF = nutrient foramen, HS = location for histological section. j SOA-MM10 right dc. From left to right, occlusal, labial, and lingual views. k SOA-MM11 left M3. From left to right, occlusal, buccal, and lingual views.
Context and geological age
All hominin fossils originated from the top of a ribbon-shaped, indurated pebbly sandstone layer (Layer II), which was deposited in a small stream channel on a volcaniclastic alluvial fan5 ~ 0.7 Ma ago. This age estimate is based on the identification of the Brunhes-Matuyama boundary15 dated at 0.773 Ma by palaeomagnetic measurements combined with a series of fission track dates on zircons in tuffaceous strata stratigraphically 16.5 m below Layer II5,6. A minimum age of 0.65 ± 0.02 Ma for Layer II is provided by a \(\ce{^40Ar}/\ce{^39Ar}\) date on single hornblende crystals from an airfall tephra (PGT-2) occurring stratigraphically at 14 m above Layer II.
Layer II, with a maximum thickness of 50 cm, overlies a reddish paleosol (Layer III) with an undulating erosional contact. A series of massive, tuffaceous clay-rich mudflow layers (Layers Ia-f) sealed off Layers II and III subsequently (Fig. 2; Supplementary Note 1). The humerus fragment SOA-MM9 was retrieved in several pieces within one week of opening Excavation 32 A in 2013, but was recognized as such only in 2015 after reconstruction in the laboratory. The specimen was damaged in the process of excavating it from the extremely compact sandstone of Layer II. A maxillary deciduous canine (dc: SOA-MM10) was excavated in 2015 at ~5 cm below the boundary between Layers I and II, while a mandibular third molar (M3: SOA-MM11) was excavated in 2016 at ~15 cm below the top of Layer II. All hominin fossils are concentrated in the upper part of Layer II, while fossils of other fauna tend to be distributed more evenly in this unit. There is evidence for fluvial transportation of the fossils prior to burial, with many (but not all) specimens fractured (apart from excavation damage), weathered and/or rounded to some extent16. However, the three hominin fossils described here show minimal/no evidence of abrasion. Taphonomic and sedimentological observations suggest that the hominin fossils were deposited during a moderate to low-energy flow regime in the stream, following a relatively brief period on the surface during which the remains were disarticulated (Supplementary Note 1). Shortly after incorporation of the fossils in the stream bed the entire stream valley was filled with a 6.5 m thick sequence of mudflows. Succeeding field excavations in 2017–2019 and 2023 have yielded no more hominin fossils from this site.Developmental age of the humerus (SOA-MM9)Fig. 2: Context of the Mata Menge hominin fossils. a Digital Elevation Map (DEM) of Flores showing the location of the So’a Basin and the cave Liang Bua. b DEM of the So’a Basin showing the location of the Mata Menge excavations. c Photo of the west baulk of the southern excavation sector (Sector 32C) in the upper fossil-bearing interval at Mata Menge. Layer III is a reddish sandy paleosol, overlain with an erosional contact by a sandy fluvial layer (Layer II). Layers II and III are covered by a series of clay rich ashy mudflow units (Layers Ia-f). Deciduous canine SOA-MM10 was recovered at 5 cm below the top of Layer II (indicated with the blue dashed rectangle; the blue rectangle is enlarged in e). Also note the large Stegodon bones resting on top of Layer II and covered by the mudflow units. d Photo of the northwest corner of the excavation in sector 32 A, taken on 7 November 2013, four weeks after the retrieval of the hominin humerus fragment SOA-MM9. The fossil was excavated from the top of Layer II, with the approximate position indicated with the dashed blue oval. e Detail of the contact between Layer I and Layer II at the spot of the deciduous canine SOA-MM10. f SOA-MM10 still partly embedded in the sandstone of Layer II. g Mata Menge excavation grid (UTM Zone 51 L) showing the 1 ×1 m quadrants excavated towards the end of the 2016 field season in gray. The positions of the hominin fossils described in this paper are indicated with green dots, those described previously9 with gray dots. Light shading represents the step trench excavated in 2010, which first revealed the presence of the Mata Menge upper fossil-bearing interval bone bed. h SOA-MM11 surrounded by its sandstone matrix when excavated in 2016. The maps (a and b) created with GeoMapApp (www.geomapapp.org) / CC BY / CC BY (Ref. 67)”.
a Digital Elevation Map (DEM) of Flores showing the location of the So’a Basin and the cave Liang Bua. b DEM of the So’a Basin showing the location of the Mata Menge excavations. c Photo of the west baulk of the southern excavation sector (Sector 32C) in the upper fossil-bearing interval at Mata Menge. Layer III is a reddish sandy paleosol, overlain with an erosional contact by a sandy fluvial layer (Layer II). Layers II and III are covered by a series of clay rich ashy mudflow units (Layers Ia-f). Deciduous canine SOA-MM10 was recovered at 5 cm below the top of Layer II (indicated with the blue dashed rectangle; the blue rectangle is enlarged in e). Also note the large Stegodon bones resting on top of Layer II and covered by the mudflow units. d Photo of the northwest corner of the excavation in sector 32 A, taken on 7 November 2013, four weeks after the retrieval of the hominin humerus fragment SOA-MM9. The fossil was excavated from the top of Layer II, with the approximate position indicated with the dashed blue oval. e Detail of the contact between Layer I and Layer II at the spot of the deciduous canine SOA-MM10. f SOA-MM10 still partly embedded in the sandstone of Layer II. g Mata Menge excavation grid (UTM Zone 51 L) showing the 1 ×1 m quadrants excavated towards the end of the 2016 field season in gray. The positions of the hominin fossils described in this paper are indicated with green dots, those described previously9 with gray dots. Light shading represents the step trench excavated in 2010, which first revealed the presence of the Mata Menge upper fossil-bearing interval bone bed. h SOA-MM11 surrounded by its sandstone matrix when excavated in 2016. The maps (a and b) created with GeoMapApp (www.geomapapp.org) / CC BY / CC BY (Ref. 67)”.
This specimen is an undistorted, distal half of the right humeral shaft that measures 88 mm in maximum preserved length (Fig. 1; Supplementary Note 2). Despite its small size, cortical bone histomorphology of SOA-MM9 clearly indicates its adult status. We examined its development stage based on age-associated increase of osteons and related structures, a method widely utilized for age estimates of extant and fossil hominins17,18,19,20,21.
Histological sections were examined for cortical samples taken at the mid-posterior shafts of SOA-MM9 (‘HS’ in Fig. 1) and from a modern human sample (N = 20, see Supplementary Data 1). To allow for regional variation in osteon formation within each bone area21, data were collected from two additional (nearby) sites in the midshaft for all the modern human (H. sapiens) specimens (Fig. 3a). In two parameters indicative of bone maturity, Osteon Population Density (OPD)21 and Haversian Canal Index (HCI), SOA-MM9 was found to exhibit distinctly greater values (OPD = 16.5, HCI = 0.85) than in any of the modern human subadult humeri (0.0–8.9 and 0.0–0.63, respectively) (Fig. 3b, Supplementary Data 1). The values for the Mata Menge humerus are also greater than the means of our modern human adult samples (13.6 and 0.78, respectively), indicating that the SOA-MM9 individual was well within adulthood at time of death. Although the external cortical surfaces of SOA-MM9 exhibit microscopic damages that might have reduced one of the marginal osteons to half (~100 microns) (Fig. 3c), such post-depositional alterations would have limited impact on our age estimation. Even if we assume surface abrasion of 200 microns, the OPD value for SOA-MM9 would drop only slightly to ~15.8. Furthermore, the dominance of secondary osteons in the outer cortex (Fig. 3c) indicates that subperiosteal bone growth during the growth period had already been terminated in this individual22.No evidence of pathology was found in SOA-MM9. Cortical bone thinning and woven bone would be pathognomic of some metabolic disorders17, but these features are not evident in SOA-MM9. The relative cortical bone thickness of SOA-MM9 (0.07: the ratio of cortical bone thickness relative to the humeral shaft circumference) (Supplementary Data 1) is almost identical to the mean for the modern human adult sample (0.069). Patients with osteogenesis imperfecta, which may lead to short stature, exhibit subnormal OPD values19, a tendency that is in opposition to the SOA-MM9 condition. Additionally, the weak but distinct development of the lateral supracondylar ridge of SOA-MM9 (Fig. 3d) indicates normal development of the extensor carpi radialis longus muscle.Fig. 3: Evidence for adulthood of the SOA-MM9 humerus. a Three sites in a cortical section of the mid-posterior humeral shaft used for the histomorphological analyzes. ‘Middle field’ is a circle drawn in the outer one-third of the midsagittal line, and the ‘lateral field’ and ‘medial field’ are corresponding circles 3 mm apart from there. b Age-related histomorphometric values for SOA-MM9 and modern humans. The data from the three sites are plotted for all the modern human specimens, whereas the data for SOA-MM9 is from the lateral field only. The dotted lines are the means of the modern human adult subsample. See Supplementary Data 1 for the original data. c Cortical section of the posterior midshaft of SOA-MM9 observed by an ultra-high-definition microscope (VHX-7000, Keyence). The original image (left) and the same image colored for intact secondary osteons (red), fragmentary osteons (orange and yellow), and the intact inner surface (blue) (right). Note the dominance of secondary osteons around the exterior surface (downside of the images). Only one section (shown here) was produced to minimize the damage to the original specimen. d Lateral supracondylar ridge of SOA-MM9 and its CT section (the arrows). Note the weak but distinct development of the ridge as a slight eversion.
a Three sites in a cortical section of the mid-posterior humeral shaft used for the histomorphological analyzes. ‘Middle field’ is a circle drawn in the outer one-third of the midsagittal line, and the ‘lateral field’ and ‘medial field’ are corresponding circles 3 mm apart from there. b Age-related histomorphometric values for SOA-MM9 and modern humans. The data from the three sites are plotted for all the modern human specimens, whereas the data for SOA-MM9 is from the lateral field only. The dotted lines are the means of the modern human adult subsample. See Supplementary Data 1 for the original data. c Cortical section of the posterior midshaft of SOA-MM9 observed by an ultra-high-definition microscope (VHX-7000, Keyence). The original image (left) and the same image colored for intact secondary osteons (red), fragmentary osteons (orange and yellow), and the intact inner surface (blue) (right). Note the dominance of secondary osteons around the exterior surface (downside of the images). Only one section (shown here) was produced to minimize the damage to the original specimen. d Lateral supracondylar ridge of SOA-MM9 and its CT section (the arrows). Note the weak but distinct development of the ridge as a slight eversion.
Humeral size
In all available dimensions of shaft diameter/circumference and length, SOA-MM9 is smaller than LB1 (H. floresiensis) and any other adult individuals of small-bodied fossil hominins (Australopithecus and H. naledi: Supplementary Data 2). Its minimum circumference (46 mm) is less than U.W. 101-283 (47.5 mm), BOU-12/1 (52 mm), and the smallest humeri in our prehistoric modern human sample (46.5 mm, N = 1050, see Supplementary Data 2). Centroid size at the ~19% level cross-section is also the smallest compared to any sampled adult specimens of Australopithecus, Paranthropus, and Homo including H. naledi and Liang Bua H. floresiensis (Fig. 4, Supplementary Data 3). The distal shaft length measured between the NF (nutrient foramen) and hOF (superior margin of the hollow leading to the olecranon fossa) points of SOA-MM9 (58 mm) is distinctly shorter than the other hominin fossils, including LB1 (64 mm) (Supplementary Data 2), although the vertical position of NF is variable in human humeri23.



The fact that the authors of the Bible got it so badly wrong, is yet more evidence that it was written by ignorant people, not an omniscient creator god, to anyone with the intellectual integrity and humility to accept that evidence trumps evidence-free opinion.
What Makes You So Special? From The Big Bang To You

How did you come to be here, now? This books takes you from the Big Bang to the evolution of modern humans and the history of human cultures, showing that science is an adventure of discovery and a source of limitless wonder, giving us richer and more rewarding appreciation of the phenomenal privilege of merely being alive and able to begin to understand it all.
Available in Hardcover, Paperback or ebook for Kindle
Ten Reasons To Lose Faith: And Why You Are Better Off Without It

This book explains why faith is a fallacy and serves no useful purpose other than providing an excuse for pretending to know things that are unknown. It also explains how losing faith liberates former sufferers from fear, delusion and the control of others, freeing them to see the world in a different light, to recognise the injustices that religions cause and to accept people for who they are, not which group they happened to be born in. A society based on atheist, Humanist principles would be a less divided, more inclusive, more peaceful society and one more appreciative of the one opportunity that life gives us to enjoy and wonder at the world we live in.
Available in Hardcover, Paperback or ebook for Kindle

No comments :
Post a Comment
Obscene, threatening or obnoxious messages, preaching, abuse and spam will be removed, as will anything by known Internet trolls and stalkers, by known sock-puppet accounts and anything not connected with the post,
A claim made without evidence can be dismissed without evidence. Remember: your opinion is not an established fact unless corroborated.