

10,000 years or so before creationism's little god created the small flat planet with a dome over it, thinking it was a universe, as described in the creation myth in the Bible, there was a mass extinction due to global climate change. In addition to the loss of the Ice Age megafauna, such as the woolly mammoth, woolly rhinoceros, cave lions, etc. over most of Eurasia and North America when temperatures rose at the end of the last glaciation, we also lost a lot of the Ice Age-adapted plants.
But, because plants tend not to fossilise so readily as the bones of large mammals, we didn't know until now, just how extensive this loss was, and more importantly, what a similar rise in temperatures is going to mean for the extant flora.
To redress this gap, a team of researchers from the Alfred Wegener Institute, Helmholtz Centre for Polar and Marine Research (AWI), Germany, have analysed DNA recovered from plant remains in the sediment of lakes in Siberia and Alaska. In doing so, they have discovered how the temperature affects the way plants interact, tending to support one another in cold weather and competing with one another in warm weather. A rise in ambient temperature meant increasing competition and loss of mutual support.
Tell me all about the flora of Siberia at the last glacial maximum, please. During the Last Glacial Maximum (LGM) (approximately 26,500 to 19,000 years ago), Siberia's flora was shaped by the harsh climatic conditions of the Ice Age. Much of the region was covered in a cold, dry steppe-tundra environment, sometimes referred to as the "Mammoth Steppe"-a biome that supported a variety of megafauna like mammoths, woolly rhinoceroses, and bison.The team's findings are published, open access, in Nature Communications and explained in a press release from AWI:
Would you like a breakdown of specific plant species that have been found in Ice Age pollen records from Siberia?
- Vegetation Types of Siberia During the LGM
Despite the extreme cold, vast ice sheets did not completely cover Siberia. The region remained an open, arid landscape dominated by hardy, cold-adapted plants.
- Steppe-Tundra Vegetation
The dominant vegetation in Siberia was a mix of steppe and tundra species, forming the Mammoth Steppe. This biome was characterized by:
- Grasses (Poaceae) - drought-resistant and adapted to cold, including genera like Festuca and Stipa.
- Sedges (Cyperaceae) - such as Carex species, important for supporting grazing herbivores.
- Forbs (herbaceous flowering plants) - including Artemisia (sagebrush) and Astragalus (milkvetch), many of which had deep root systems to access nutrients in permafrost soils.
- Mosses and lichens - found in patches where conditions were too extreme for other vegetation.
- Shrub Tundra
In some areas, particularly southern and eastern Siberia, shrubs formed a patchy part of the landscape, including:
- Dwarf birches (Betula nana) - common in tundra environments, with small, resilient leaves.
- Willows (Salix spp.) - creeping and low-growing, adapted to cold climates.
- Arctic blueberries (Vaccinium uliginosum) - and other heathland plants, though limited in distribution.
- Sparse Coniferous Forests
While true taiga forests (dominated by larches, spruces, and pines) were limited due to the arid and frigid conditions, some conifers persisted in more sheltered, southern locations:
- Larch (Larix sibirica) - one of the few cold-resistant trees that could tolerate permafrost conditions.
- Spruce (Picea obovata) - possibly found in refugia (isolated areas where trees survived despite harsh conditions).
- Adaptations of Siberian Flora to Ice Age Conditions
Plants in LGM Siberia had to withstand freezing temperatures, low precipitation, and nutrient-poor soils. Their adaptations included:
- Perennial life cycles - many plants lived multiple years, avoiding the need for rapid seasonal growth.
- Deep root systems - helped access water locked in permafrost.
- Low, compact growth forms - protected plants from wind and cold.
- C4 photosynthesis in grasses - some steppe grasses may have used this adaptation to maximize water efficiency.
- The Decline of the Mammoth Steppe
After the LGM, as the climate warmed, Siberia saw a transition from the steppe-tundra biome to taiga forests in the north and wet tundra in the Arctic. This shift contributed to the decline of megafauna like woolly mammoths, which relied on the rich grasslands of the Ice Age steppes.
Yes, please.
Pollen records and plant macrofossils from Siberia during the Last Glacial Maximum (LGM) provide a detailed picture of the vegetation that existed in the region. These records come from lake sediments, peat deposits, and permafrost-preserved soils. Below is a breakdown of some of the most important plant groups and species identified.
- Dominant Plant Families and Genera Found in Ice Age Pollen Records
- Grasses and Sedges (Poaceae, Cyperaceae) – The Foundation of the Mammoth Steppe
Pollen evidence: Grasses and sedges were dominant across Siberia, indicating an open, arid environment.
- Poaceae (Grass family)
- Festuca (Fescue) – A common cold-resistant grass.
- Stipa (Feathergrass) – Adapted to dry, steppe-like conditions.
- Poa (Bluegrass) – Found in tundra and steppe environments.
- Cyperaceae (Sedge family)
- Carex (Sedges) – Important for soil stabilization and animal forage.
- Eriophorum (Cotton grass) – Often found in wetter tundra patches.
- Forbs (Herbaceous Flowering Plants) – Drought and Cold-Resistant Plants
Pollen evidence: The prevalence of Artemisia and other drought-resistant forbs indicates an open, steppe-like ecosystem.
- Asteraceae (Daisy family)
- Artemisia (Sagebrush, Wormwood) – One of the most abundant plants in pollen records, thriving in dry, steppe-like conditions.
- Saussurea – A high-altitude genus adapted to harsh climates.
- Fabaceae (Legume family)
- Astragalus (Milkvetch) – Nitrogen-fixing plant, enriching the soil.
- Caryophyllaceae (Carnation family)
- Silene (Campion) – Found in tundra and alpine environments.
- Brassicaceae (Mustard family)
- Draba (Whitlowgrass) – A genus adapted to periglacial conditions.
- Shrubs and Dwarf Trees – Surviving in the Cold
Pollen evidence: While steppe vegetation dominated, these shrubs and occasional trees indicate small patches of tundra and taiga elements.
- Betulaceae (Birch family)
- Betula nana (Dwarf birch) – A common low shrub in tundra regions.
- Salicaceae (Willow family)
- Salix (Willows) – Often low-growing, hugging the ground to avoid wind exposure.
- Ericaceae (Heath family)
- Vaccinium (Bilberries, Lingonberries) – Found in tundra and boreal settings.
- Pinaceae (Pine family)
- Larix sibirica (Siberian larch) – One of the only conifers capable of surviving permafrost conditions in refugia.
- Picea obovata (Siberian spruce) – Present in some isolated areas.
- Regional Variations in Siberian Ice Age Flora
- Western Siberia: Slightly wetter, with more sedges (Carex), some larch trees (Larix), and isolated patches of spruce (Picea).
- Eastern Siberia (Yakutia, Chukotka): More arid, dominated by Artemisia, grasses (Festuca, Stipa), and scattered dwarf birch (Betula nana).
- Far Northern Siberia (Near Arctic Ocean): Mostly non-vascular plants (mosses and lichens), along with occasional willows (Salix).
- Changes Over Time
- Before the LGM (~40,000 years ago): Slightly warmer and wetter, with more shrubs and conifers.
- During the LGM (26,500–19,000 years ago): Cold and dry, dominated by steppe grasses and Artemisia.
- After the LGM (~14,000 years ago onward): Climate warmed, leading to the expansion of boreal forests and the decline of steppe vegetation.
Conclusion
The pollen record strongly supports the idea that LGM Siberia was covered primarily by a cold steppe-tundra ecosystem, rich in grasses, Artemisia, and sedges, with some shrubs in milder microclimates. This vegetation supported Ice Age megafauna like woolly mammoths, reindeer, and steppe bison. As the climate warmed, the dominance of these plants declined, giving way to modern taiga forests.
Global warming and mass extinctions: What we can learn from plants from the last ice age
New analysis methods, applied to ancient plant DNA, reveal how hard-hit plants were and are by global warming
Global warming is producing a rapid loss of plant species – according to estimates, roughly 600 plant species have died out since 1750 – twice the number of animal species lost. But which species are hit hardest? And how does altered biodiversity actually affect interactions between plants? Experts from the Alfred Wegener Institute have tackled these questions and, in two recent studies, presented the answers they found buried in the past: using fragments of plant genetic material (DNA) deposited in lake sediments, they were able to gain new insights into how the composition of flora changed 15,000 to 11,000 years ago during the warming at the end of the last ice age, which is considered to be the last major mass extinction event before today. This comparison can offer an inkling of what might await us in the future. The researchers have just published their findings in the journal Nature Communications.
Everyone knows that the woolly mammoth went extinct, but virtually no-one mentions the plants that were lost at the end of the last ice age. Until recently, we lacked suitable methods for investigating the extinction of plant species in detail.
Using cutting-edge methods, we analysed old DNA from sediment cores taken from lakes in Alaska and Siberia, which allowed us to reconstruct the changes in vegetation in these regions.
Professor Ulrike Herzschuh, lead author
Alfred Wegener Institute
Helmholtz Centre for Polar and Marine Research (AWI).
In terms of fossil plant remains, mainly pollen was used, which doesn’t allow individual species to be identified and therefore offers no evidence of which species have died out. The cores contain fragmented DNA from deposited plant biomass from the past 30,000 years, which the experts enriched, sequenced, and compared with databases for identification purposes at special-purpose labs for old DNA.
Temperature can change how plants interact Floating drilling platform for collecting sediment cores from lakes, Alaska.Photo: Alfred-Wegener-Institut / Weihan Jia>
Floating drilling platform for collecting sediment cores from lakes, Alaska.Photo: Alfred-Wegener-Institut / Weihan Jia>We’ve now been able to determine in detail when and where species appeared and disappeared in Alaska and Siberia. Our research shows that the composition of plant species changed substantially at the end of the last ice age, and that this was accompanied by fundamental changes in the ecological conditions.
Professor Ulrike Herzschuh.
The researchers identified a connection between temperature and plant-to-plant interactions: in cold climate periods, plant species support one another, while they mainly compete during warm periods.
In the DNA from the lake sediments, we found e.g. many cushion plants, which most likely supported the expansion of other species by forming sheltered habitats.
Professor Ulrike Herzschuh.
This has effects on both biodiversity and richness range size. In a warmer climate, woody plant species dominate: 'Today, we see that plant diversity declines due to the migration of trees and shrubs into tundra regions, whereas during cold periods, higher plant diversity prevailed.
What does that tell us about vegetation changes in the high latitudes, where cushion plants still play a pivotal role today? In today’s Arctic, this supportive quality could actually threaten their own survival.
Since the warming of the Arctic has already progressed quite far, woody plants can survive even in the high latitudes. The cushion plants could facilitate their spreading, hastening their own extinction in the process.
Professor Ulrike Herzschuh.
Which plant species are particularly at risk?
 Extraction of ancient DNA from lake sediment samples in the paleogenetics laboratory at the AWI in PotsdamPhoto: Alfred-Wegener-Institut / Janine Klimke.
Extraction of ancient DNA from lake sediment samples in the paleogenetics laboratory at the AWI in PotsdamPhoto: Alfred-Wegener-Institut / Janine Klimke.
The end of the last ice age also caused some types of vegetation to disappear entirely – as the experts were able to confirm using their new methods. Take the mammoth steppe, for example: during the last ice age, this type of vegetation spread across the Northern Hemisphere, only to die out during the transition to the current age. In this regard, identifying the extinct plant species was especially challenging.
To identify the species that no longer existed, we had to use a trick. [Normally, species are identified on the basis of DNA fragments, which are compared with the entries in genetic databases. But these databases include information on today’s plants, not on extinct species]. We examined all the DNA fragments from our cores and then used statistical models to filter out those with unmistakeable similarities to modern plants, step by step.
Professor Ulrike Herzschuh.
This also allowed the experts to determine which species could be at the greatest risk of extinction in a warming world: grasses and shrubs are at a higher risk of disappearing than woody plant species, which can spread further when temperatures rise. In addition, species in regions with high biodiversity are more often at risk than are less “special” species. One surprising finding: the extinction rate was at its highest at the beginning of the current warm phase – often with a delay of several thousand years after the actual environmental changes. “That means the full impacts of today’s human activities might not become apparent until the distant future.”
Relevance for today’s Arctic
The results of the two studies offer fundamental insights into how environmental changes in connection with warming affect biodiversity, and which mechanisms are central in this regard. As such, for the first time the experts were able to determine extinction rates for plants, which can now be used as reference data to better assess the ongoing changes in Arctic ecosystems.
Our studies show how important it is to understand biodiversity and ecological interactions, also in the long term, in order to better predict the impacts of climate change. Using the information locked in old DNA from sediments, we can gain the fundamental knowledge needed to do so.
Professor Ulrike Herzschuh.
Further information:Original publication
Courtin, J., Stoof-Leichsenring, K. R., Lisovski, S., Liu, Y., Alsos, I. G., Biskaborn, B., K., Diekmann, B., Melles, M., Wagner B., Pestryakova, L., Russel, J., Huang, Y., Herzschuh, U. (2025). Potential plant extinctions with the loss of the Pleistocene mammoth steppe. Nature Communications (doi:10.1038/s41467-024-55542-x)
Liu, Y., Lisovski, S., Courtin, J., Stoof-Leichsenring, K.R., Herzschuh, U. (2025)
Plant interactions associated with a directional shift in the richness range size relationship during the Glacial-Holocene transition in the Arctic. Nature Communications (doi:10.1038/s41467-025-56176-3)


AbstractThe general finding of this research is that extinctions are not triggered by a failure to adapt to climate change per se or to speciation triggered by that change, but due to changes in the distribution of major plants within the ecosystem. As the 'cushion' plants such as mosses and saxifrages retreat, so the protection they afforded to other plants disappears. Meanwhile, increased tree growth causes a competition for light so plants may be simultaneously deprived of protection and exposed to competition.
A nearly ubiquitous negative relationship between taxonomic richness and mean range-size (average area of taxa) is observed across space. However, the complexity of the mechanism limits its applicability for conservation or range prediction. We explore whether the relationship holds over time, and whether plant speciation, environmental heterogeneity, or plant interactions are major factors of the relationship within northeast Siberia and Alaska. By analysing sedimentary ancient DNA from seven lakes, we reconstruct plant richness, biotic environmental heterogeneity, and mean range-size over the last 30,000 years. We find positive richness to range-size relationships during the glacial period, shifting to negative during the interglacial period. Our results indicate neither speciation nor environmental heterogeneity is the principal driver. Network analyses show more positive interactions during the glacial period, which may contribute to positive richness to range-size relationships. Conversely, in the interglacial environment, negative interactions may result in negative relationships. Our findings suggest potential susceptibility to invasion but conservation advantages in far northern tundra given their positive interactions.
Introduction
As a universal characteristic of every species, geographic range-size is a strong predictor of species expansion and vulnerability to extinction, and can be regarded as an indicator for species conservation status. A negative relationship between richness and mean range-size is nearly ubiquitously observed across space for various taxonomic groups1,2,3, yet the exact mechanism of the richness to mean range-size relationship is still poorly understood. Furthermore, it remains unclear whether the spatial relationship holds true in the temporal domain. It is of particular interest whether the observed species richness decline in the context of global change4 is associated with an increase in mean range-size. Alternatively, do changes in species range-size result in richness adjustments? Answering these questions requires the knowledge to exploit the richness to range-size relationship for predictions, for example, in the context of conservation efforts5,6.
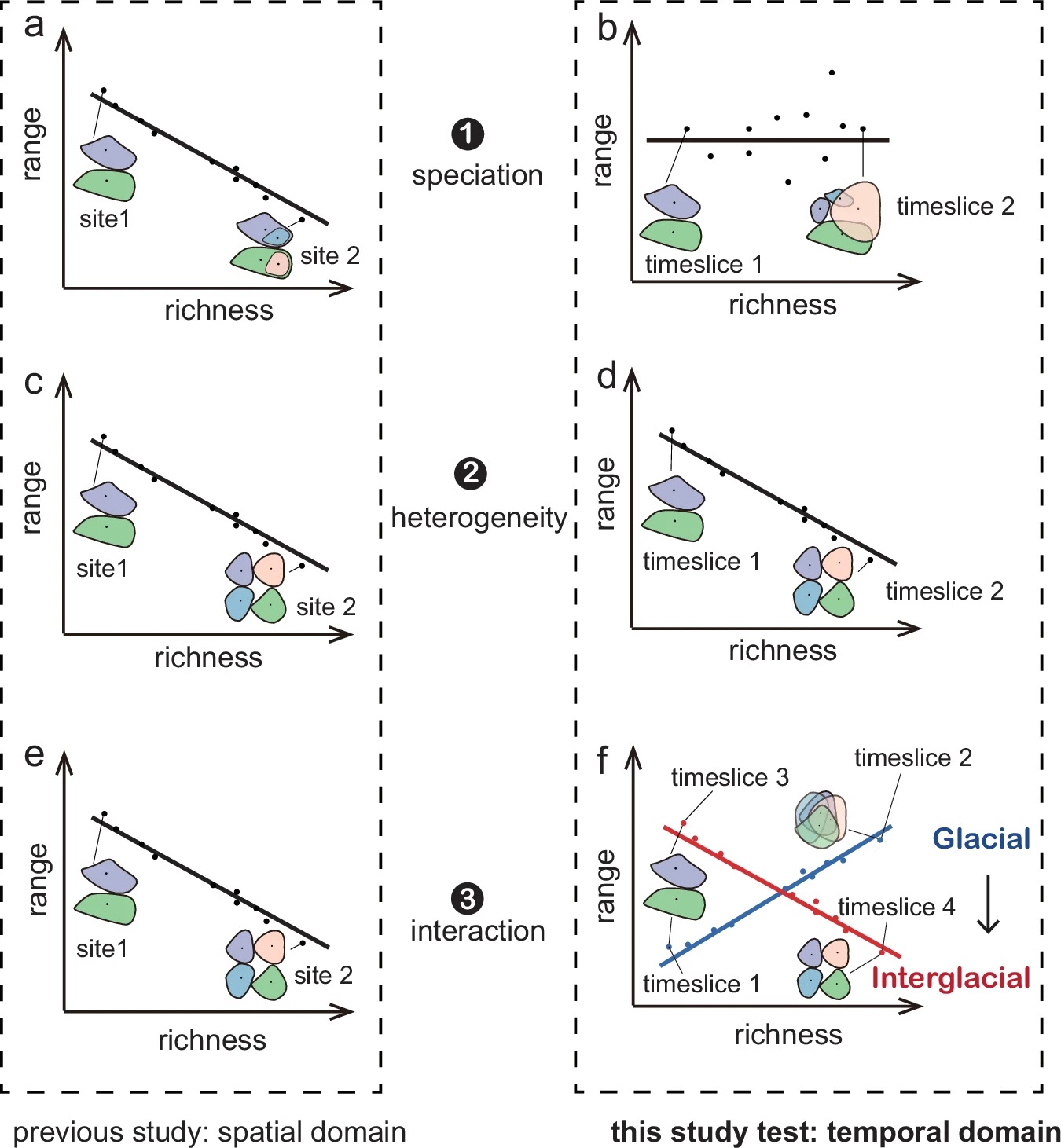
Based on previous research, three main hypotheses are proposed to explain the widely observed negative richness to range-size relationship. First, long-term (millions of years) temperature-related speciation and species extinction may result in negative richness to mean range-size relationships in the spatial domain7,8. Higher temperatures increase metabolic rates and promote speciation rates9. Within a region with high speciation rates, new species are initially constrained within a small range, resulting in regions with high species richness and smaller range-sizes8,10 as exemplified by the equatorial region11. In contrast, low temperatures and recurrent extensive glaciation in high-latitude regions have resulted in less speciation and increased the risk of regional species extinctions12. Thus, variation in speciation and extinction rates along the temperature gradient results in a negative richness to range-size relationship in space (Fig. 1a)7,8. However, this hypothesis applies to the global pattern; it is not solely attributed to temperature differences across space but also to the temporal differences in species accumulation and extinction over millions of years. Within a specific region and over a millennial time scale, the impact of speciation and extinction may be limited. Hence, to shift the perspective to a temporal domain over millennia, richness and range changes are not driven by speciation. Moreover, without considering extinction, there is no expected relationship between richness and range-size across time (Fig. 1b).
Second, environmental heterogeneity can result in a negative richness to mean range-size relationship13,14. Heterogeneous environments, for example complex forest ecosystems, may promote the colonisation of specialised taxa that can coexist15,16. This leads to a high richness of taxa with narrow ecological niches and limited geographical ranges13, resulting in a negative richness to range-size relationship along spatial heterogeneity gradients (Fig. 1c). Accordingly, in the temporal domain, a stable negative richness to mean range size relationship will be expected (Fig. 1d). For example, the diversity decline related to the expansion of generalist taxa with a large distribution range can be explained by human landscape homogenisation17,18.Fig. 1: Evaluating three hypotheses of how species richness relates to range-size over space (previous studies3,7,8,13,14) and time (this study), using a hypothetical richness increase scenario. Coloured polygons represent the distribution range of different plant taxa. Under the speciation hypothesis, regions experiencing high speciation rates over millions of years generate taxa with a constrained range, resulting in a negative richness to range-size relationship (a), in the temporal domain, no speciation occurs on millennial time scales, neither during the glacial period nor the interglacial period. The richness increases are not driven by speciation, and range-size remains stable or changes without a discernible pattern, thus, there is no richness to range-size relationship (b). Under the environmental heterogeneity hypothesis, the temporal domain pattern (d) mirrors the spatial domain pattern (c). Spanning both glacial and interglacial periods, heterogeneous environments favour a high richness of taxa with narrow ecological niches and limited geographic ranges, leading to a stable negative richness to range-size relationship (d). Under the plant interaction hypothesis, widespread negative richness to range-size relationships are observed in the spatial domain, attributed to negative interactions (e). Unlike the spatial domain result, in the temporal domain, a positive interaction during the glacial period expands taxa range-sizes (f), resulting in a positive richness to range-size relationship; while a negative interaction during the interglacial period constrains the range-sizes of the taxa, resulting in a negative relationship. The relationship shifts from positive during the glacial towards negative during the interglacial (f).
Coloured polygons represent the distribution range of different plant taxa. Under the speciation hypothesis, regions experiencing high speciation rates over millions of years generate taxa with a constrained range, resulting in a negative richness to range-size relationship (a), in the temporal domain, no speciation occurs on millennial time scales, neither during the glacial period nor the interglacial period. The richness increases are not driven by speciation, and range-size remains stable or changes without a discernible pattern, thus, there is no richness to range-size relationship (b). Under the environmental heterogeneity hypothesis, the temporal domain pattern (d) mirrors the spatial domain pattern (c). Spanning both glacial and interglacial periods, heterogeneous environments favour a high richness of taxa with narrow ecological niches and limited geographic ranges, leading to a stable negative richness to range-size relationship (d). Under the plant interaction hypothesis, widespread negative richness to range-size relationships are observed in the spatial domain, attributed to negative interactions (e). Unlike the spatial domain result, in the temporal domain, a positive interaction during the glacial period expands taxa range-sizes (f), resulting in a positive richness to range-size relationship; while a negative interaction during the interglacial period constrains the range-sizes of the taxa, resulting in a negative relationship. The relationship shifts from positive during the glacial towards negative during the interglacial (f).
Third, biotic interactions among taxa may drive richness to range-size relationships3. However, there is a lack of systematic investigations along spatial gradients. The stress-gradient hypothesis19,20 suggests that in less stressful environments, negative interactions are promoted, while in stressful environments, positive interactions should be more common. In line with the stress-gradient hypothesis, it may be hypothesised that in less stressful environments, high richness can lead to increased competition and predation (negative interactions), limiting growth rate21 and reducing range-sizes3, resulting in a negative richness to-range-size relationship. In contrast, environmental stress can promote positive interactions among taxa allowing for mutually wide ranges19,20,22,23, which would be reflected by a positive richness to range-size relationship. However, positive richness to range-size relationships are rarely detected in space, which may be because other factors obscure the relationship when multiple drivers are correlated. In the temporal domain, an environmental stress release may lead to a shift from a positive to a negative richness to range-size relationship (Fig. 1). For example, this shift might occur when transitioning from the stressful tundra environment during the glacial period to the less stressful taiga environment in the Holocene24,25,26 (Fig. 1f).
To disentangle the potential factors causing richness to range-size relationships, conducting studies from a temporal perspective within confined geographical regions and appropriate palaeoenvironmental settings is essential. For this purpose, northeast Siberia plus Alaska is a suitable study area. Because of polar amplification, this high-latitude area has experienced strong changes in temperature on millennial time scales23, mirrored by ecological turnover along with treeline translocation27. However, unlike other climate-sensitive high-latitude regions, this area was not covered by ice shields during the last glacial period28,29. As such, lagged species assembly effects in the course of new habitat formation after deglaciation30 are minimal due to the potential for regional recruitment. The area also provides high-quality lake sedimentary archives reaching back to the Last Glacial Maximum (LGM; 21 ka BP) and beyond. Furthermore, previous studies from the area have yielded high-quality sedimentary ancient DNA (sedaDNA) records. Compared to traditional pollen analysis, sedaDNA can identify past plants to a higher taxonomic resolution, which allows the reconstruction of past plant richness and distribution changes at a reasonable taxonomic level31.
Here, we use sedaDNA-based plant community reconstructions of the last 30,000 years to investigate richness to mean range-size relationships in the temporal domain within northeast Siberia plus Alaska (Fig. 1). Our straightforward study design will test the following hypotheses. (1) If millions of years of speciation and species loss are the primary factors, when applying this pattern to millennial studies, especially after eliminating the potential impact of extinction, an insignificant richness to range-size relationship would be expected through time. In contrast, if (2) heterogeneity dominates, which can change on millennial time scales, we expect that higher species richness is consistently related to smaller range-size over time. Alternatively, (3) along with postglacial forest expansion, interactions might have changed from a system that is dominated by mutualistic (positive) interactions to systems dominated by competitive (negative) interactions, which should be reflected by a shift from a positive to a negative richness to mean range-size relationship. Overall, our study aims to disentangle the potential factors, providing insight into temporal plant richness and mean range-size relationships to inform future conservation and protection.
Liu, Y., Lisovski, S., Courtin, J. et al.
Plant interactions associated with a directional shift in the richness range size relationship during the Glacial-Holocene transition in the Arctic Nat Commun 16, 1128 (2025). https://doi.org/10.1038/s41467-025-56176-3
Copyright: © 2024 The authors.
Published by Springer Nature Ltd. Open access.
Reprinted under a Creative Commons Attribution 4.0 International license (CC BY 4.0)
The problem for creationists however, is not that this sort of change in fauna can be predicted from the Theory of Evolution but is incomprehensible as the work of an intelligent designer, it is the fact that this change took place thousands of years before they believe Earth and all life on it were magically created out of nothing, and that the evidence for the change still exists, in the sediment in lakes, where ancient plant matter and ancient DNA can still be found.








No comments :
Post a Comment
Obscene, threatening or obnoxious messages, preaching, abuse and spam will be removed, as will anything by known Internet trolls and stalkers, by known sock-puppet accounts and anything not connected with the post,
A claim made without evidence can be dismissed without evidence. Remember: your opinion is not an established fact unless corroborated.