
Entrapment of an elephant in the quicksand of layer 5 of the upper archaeological level of the late Early Pleistocene site of Fuente Nueva-3.

A team of archaeologists from Malaga University have uncovered the earliest evidence of ancient hominin activity in Europe - at 1.4 million years ago.
The evidence from the Fuente Nueva-3 archaeological site in Orce, southeastern Spain shows that hominins scavenged alongside giant hyenas for animals which became stuck in a quicksand trap in what is now the Guadix-Baza Depression.
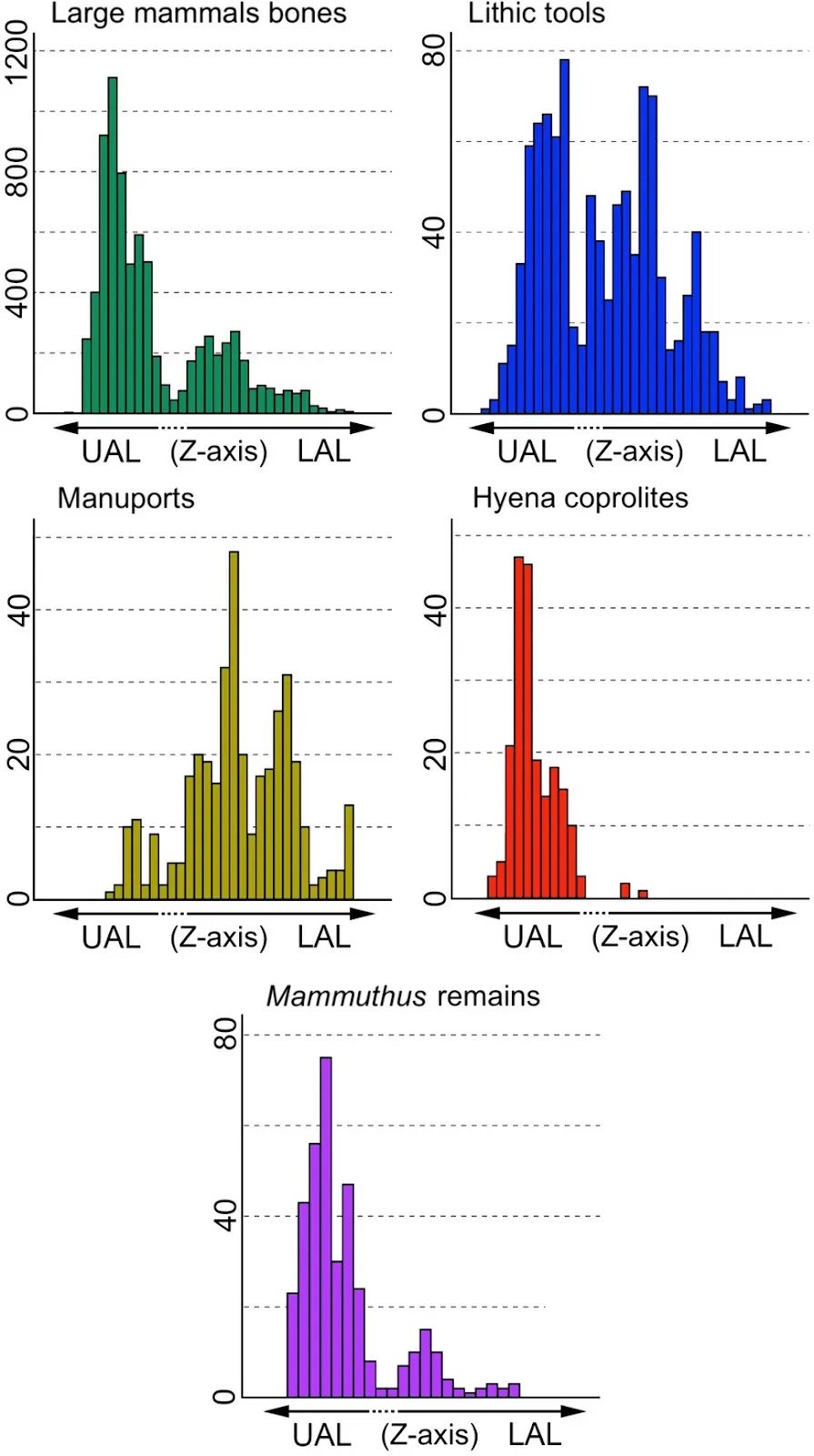
The team included Professor of Paleontology Paul Palmqvist and Professor of Stratigraphy and Paleontology María Patrocinio Espigares. They showed that the site consisted of two distinct archaeological levels, an upper (UAL) and a lower (LAL) level, the latter of which contains a high density of manuports, while the UAL contains remains of megafauna such as mammoths, Mammuthus meridionalis, and many hyena coprolites, showing significant scavenging activity
What information do you have on the Fuente Nueva-3 archaeological site in the Guadix-Baza Depression in Spain? The Fuente Nueva-3 archaeological site is located in the Guadix-Baza Depression in southern Spain. This site is of significant importance due to its contribution to the understanding of early human habitation in Europe. Here are some key points about Fuente Nueva-3:The team's findings are published in the Journal of Iberian Geology and are explained in a press release from Malaga University (reprinted in MirageNews.com):
In summary, Fuente Nueva-3 is a key archaeological site that offers valuable insights into the early human occupation of Europe, their technological capabilities, and their interactions with the environment during the Early Pleistocene.
- Geographical Context:
- The Guadix-Baza Depression is a sedimentary basin in the Betic Cordillera of southern Spain, characterized by a semi-arid climate. It contains rich Pleistocene deposits, making it an important area for paleoanthropological and archaeological studies.
- Age and Chronology:
- Fuente Nueva-3 dates back to the Early Pleistocene, approximately 1.4 million years ago. This makes it one of the oldest archaeological sites in Europe, providing evidence of early human presence on the continent.
- Archaeological Findings:
- The site has yielded numerous lithic artifacts, including tools made from local stone materials. These tools exhibit the characteristic features of Mode 1 technology, also known as Oldowan, which includes simple flake tools and choppers.
- In addition to stone tools, Fuente Nueva-3 has produced a rich assemblage of faunal remains. These include bones of large mammals such as elephants, rhinoceroses, and various species of deer and horses, indicating that early humans at the site engaged in activities like hunting or scavenging.
- Paleoenvironmental Evidence:
- The faunal assemblage and sedimentary context suggest a varied landscape with open grasslands, water bodies, and some wooded areas. This environment would have supported a diverse range of megafauna, providing ample opportunities for early humans to exploit for food resources.
- Significance for Human Evolution:
- The findings at Fuente Nueva-3 are crucial for understanding the dispersal of early humans into Europe. The site's early date, combined with the technological and faunal evidence, supports the hypothesis that hominins left Africa and adapted to a range of environments across Eurasia earlier than previously thought.
- It also provides insights into the behavior and adaptive strategies of these early hominins, particularly in terms of their tool-making abilities and subsistence practices.
- Research and Excavations:
- Ongoing excavations and research at Fuente Nueva-3 have involved multidisciplinary teams, including archaeologists, paleontologists, and geologists. These efforts have led to a more comprehensive understanding of the site's stratigraphy, chronology, and the context of the artifacts and fossils found there.
UMA Scientists Discover Ancient Quicksand Trap in Orce
Scientists at the University of Malaga have shown in an unprecedented way how the so-called 'elephant graveyard' of the Early Pleistocene archaeological site of Orce – name given due to the amount of remains of the extinct elephant species Mammuthus meridionalis that it contained – hid a natural trap in quicksands.
The site of Fuente Nueva 3 (FN3), located in the northeastern margin of the Guadix-Baza Depression (Granada), is one of the deposits of Orce that contains some of the earliest evidence of human presence in Western Europe – consisting of lithic assemblages, that is, stones carved by our ancestors –, that is dated to one million four hundred thousand years ago.
Likewise, this archaeological site preserves manuports –unmodified stones used as percussion tools to fracture bones and access the marrow and, perhaps, also used as throwing weapons to scare away hyenas– and abundant fossils of large mammals, some of which preserve anthropogenic marks related to skinning, butchering and marrow processing. And also bones with tooth marks caused by scavenging carnivores.
The results of this pioneering research have recently been published in Iberian Journal of Geology –of the prestigious publishing house Springer-Nature.
Two archaeological levels
According to this study, co-led by Full Professor of Paleontology at the UMA Paul Palmqvist and Professor of Stratigraphy and Paleontology at the UMA María Patrocinio Espigares, the fertile layers of this site have two different archaeological levels: lower (LAL) and upper (UAL). Both levels preserve abundant skeletal remains and lithic tools. However, while the first one shows a high density of manuports, which suggests that hominin activity was more intense at this level, the second one preserves many remains of megaherbivores, particularly, the extinct elephants Mammuthus meridionalis, which points to a greater involvement of the giant hyenas.
Thus, these scientists have analyzed the statistical differences in the composition of the faunal assemblages preserved in these two levels and the sedimentology, specifically the particle size in the fertile layers of both levels. The latter is a key aspect in the research, as it shows a predominance of silts and clays in layers 2-3 of the lower level, and fine and very fine sands in layer 5 of the upper level. "These fine sand sediments, deposited close to the paleolake that was in the region, would also contain slightly saline water, a mixture that explains that they could have worked as quicksand, where larger animals were trapped," say the UMA scientists.
Food for scavengers
Therefore, experts point out that the latter level may be interpreted as a natural quicksand trap in which megaherbivores were trapped due to the elevated weight of their limbs, and their half-sunken carcasses attracted the scavengers, both hyenas and humans, which fed on these and left their lithic assemblages and coprolites –fossilized excrement of hyenas– as evidence of their presence.
The new finding discovered by UMA researchers represents a "very important milestone to gain more knowledge about the subsistence strategies of our ancestors, the first Europeans, and their competition with the large carrion hyenas for access to these meat resources, since it is the first time a natural trap with these characteristics has been described in a fossil deposit of special interest to human evolution."
Carrying out more detailed studies differentiating the upper and lower archaeological levels, as well as characterizing other important sites in the Orce region, such as Barranco León, which also contributes evidence of human presence, are the next steps planned by this scientific team of the UMA, in which researchers from the Complutense University of Madrid and Tarragona also participate.
More than a decade of UMA research in Orce
The University of Malaga has been researching the site of Fuente Nueva 3 for several decades. The first publication of the UMA that highlighted the significance of this place, in 2013, described a partial elephant skeleton surrounded in part by flint flakes and coprolites, attesting to the competition between these two super-scavengers for the carcass resources.
Bibliography:Palmqvist, P., Campaña, I., Granados, A. et al. (2024). The late Early Pleistocene site of Fuente Nueva-3 (Guadix-Baza Depression, SE Spain): a hyena latrine developed on a quicksand trap for megaherbivores? J Iber Geol. https://doi.org/10.1007/s41513-024-00241-1
AbstractAgain, we see evidence that the authors of Genesis were ignorant of the world outside their immediate vicinity and time, comprehensively refuting the idea that it was written, dictated or inspired by a supernatural, omniscient creator god. It is a work of imagination by people with limited horizons, limited knowledge, minimal understanding of the world and little in the way of science, and so limited imagination, which is why most of it is hopelessly wrong.
The late Early Pleistocene archaeological site of Fuente Nueva-3 (FN3), which lies in the Guadix-Baza Depression (SE Spain) and is dated to ~ 1.4 Ma, contributes some of the oldest evidence of hominin presence in Western Europe, including a huge tool assemblage of Oldowan tradition, manuports (i.e., unmodified stones used as percussion tools) and abundant fossils of large mammals, some of which preserve anthropogenic marks related to defleshing, butchering and marrow processing. In addition, there are bones with tooth marks produced by scavenging carnivores. The fertile layers of the FN3 section have been grouped in a lower archaeological level (LAL) and an upper archaeological level (UAL). Both levels preserve abundant skeletal remains and lithic tools. However, the LAL shows a high density of manuports, which suggests that hominin activity was more intense at this level, while the UAL preserves many remains of megaherbivores, particularly proboscideans (Mammuthus meridionalis), and almost all coprolites unearthed from the site, which points to a greater involvement of the giant hyenas (Pachycrocuta brevirostris). In this paper, we (i) test for statistical differences in the composition of the faunal assemblages preserved in the UAL and LAL; and (ii) analyze particle size in the fertile layers of both archaeological levels. Our results show that megaherbivores are comparatively overrepresented in the UAL, specially by young elephants, while other medium-to-large and large-sized ungulates, particularly equids, are more abundant in the LAL, showing a predominance of adult individuals. Concerning the sedimentology, layers 2–3 of the LAL show a predominance of silts and clays, with sands representing a minor fraction. In contrast, layer 5 of the UAL is composed of two-thirds of fine and very fine sands, with the remaining third consisting of silts and clays. These data and the paleoenvironmental reconstruction of layer 5 suggest that it may be interpreted as a paleo-quicksand in which megaherbivores were trapped due to the elevated weight per unit area supported by their feet, and their half-sunken carcasses attracted the scavengers, particularly the hyenas, which fed on these carcasses and defecated in their surroundings.
1 Introduction and background
The late Early Pleistocene archaeo-paleontological site of Fuente Nueva-3 (FN3) lies in the north-eastern sector of the Guadix-Baza Depression (SE Spain, Fig. 1A–B), which is the largest intramontane basin of the Betic Cordillera (García-Aguilar and Palmqvist 2011; García-Tortosa et al. 2011.1; Palmqvist et al. 2022a). This inland basin extends over an area of ~ 4000 km2 and is surrounded by the highest reliefs of the Alpine-Betic orogeny, which rise up to an altitude of ~ 3482 m above mean sea level (AMSL). During the Miocene, the sedimentary depression was connected to the Mediterranean Sea by the Almanzora Corridor and to the Atlantic Ocean by the Guadalquivir Basin. Both corridors were closed between the end of the Tortonian and the Lower Messinian, which led to a change from exorheic conditions to continental sedimentation in an endorheic basin (Soria et al. 1999; Hüsing et al. 2010). Isostatic uplifting, with an average uplift rate of ~ 200 m/Ma as estimated from Late Neogene coastal marine conglomerates and coral reefs (Braga et al. 2003), resulted in an elevation of the uppermost sedimentary level of the basin (i.e., the glacis geomorphological surface) to ~ 1000 m AMSL.The Guadix-Baza Depression developed a network of endorheic drainage during most of the Pliocene and Pleistocene, being subject to intense tectonic subsidence in relation to the surrounding mountain ranges. This led to the accumulation of a ~ 550 m thick sedimentary record composed of lacustrine, alluvial and fluvial deposits (limestones, gypsum, marls, shales, sands, and conglomerates) Tectonically induced hydrothermal activity in the basin provided a mild and productive environment for the terrestrial fauna (García-Aguilar et al. 2014, 2015; Palmqvist et al. 2022a). This favored the development of rich and diverse paleocommunities of mammals through the Plio-Pleistocene, whose fossil remains were abundantly preserved in many paleontological and archaeological localities (Maldonado-Garrido et al. 2017; Martínez-Navarro et al. 2018; Palmqvist et al. 2022a). The Orce area (Fig. 1B–C), a satellite basin in the north-eastern sector of the depression with an extent of ~ 170 km2, preserves a unique fossil record of the late Early Pleistocene (Late Villafranchian) fauna, including two archaeological sites dated to ~ 1.4 Ma that mark the earliest presence of hominins in Western Europe, Barranco León-D (BLD) and FN3 (Martínez-Navarro et al. 1997, 2014.1; Espigares et al. 2013, 2019, 2023; Toro-Moyano et al. 2013.1; Palmqvist et al. 2016, 2022a, 2023.1a).Fig. 1 A Geological context of the Baza Basin (SE Spain) within the Betic Cordillera (area marked by the rectangle). B Geologic map of the Baza Basin. 1- Substrate of the External Zones; 2- Substrate of the Internal Zones; 3- Tortonian, marine facies; 4- Plio-Pleistocene, alluvial facies; 5- Ruscinian-Lower Villafranchian, mainly lacustrine facies; 6- Lower and Middle Villafranchian, distal fluvial facies (sandstones and lutites); 7- Lower and Middle Villafranchian, marly-evaporitic lacustrine facies; 8- Late Lower Pleistocene lacustrine and fluvio-lacustrine facies; 9- Late Lower and Middle Pleistocene alluvial and lacustrine facies; 10- Glacis surface; 11- Recent alluvial; 12- Recent fluvial terraces and deposits; 13- Tectonic fractures (in discontinuous trace, inferred faults). C Geographic and geologic context of the Orce-Venta Micena sector in the Baza Basin. 1- Dolomites and limestones of the Subbetic domain (External Zones); 2- Red alluvial conglomerates; 3- Yellow lacustrine marls; 4- Lacustrine limestones; 5- Lacustrine marls, sands and gypsum; 6- Red and grey fluvial and lacustrine sands and clays; 7- Lacustrine marls and limestones; 8- Alluvial conglomerates; 9- Alluvial-deltaic conglomerates and sands, and lacustrine marls; 10- Glacis surface; 11- Recent fluvial gravels.
A Geological context of the Baza Basin (SE Spain) within the Betic Cordillera (area marked by the rectangle). B Geologic map of the Baza Basin. 1- Substrate of the External Zones; 2- Substrate of the Internal Zones; 3- Tortonian, marine facies; 4- Plio-Pleistocene, alluvial facies; 5- Ruscinian-Lower Villafranchian, mainly lacustrine facies; 6- Lower and Middle Villafranchian, distal fluvial facies (sandstones and lutites); 7- Lower and Middle Villafranchian, marly-evaporitic lacustrine facies; 8- Late Lower Pleistocene lacustrine and fluvio-lacustrine facies; 9- Late Lower and Middle Pleistocene alluvial and lacustrine facies; 10- Glacis surface; 11- Recent alluvial; 12- Recent fluvial terraces and deposits; 13- Tectonic fractures (in discontinuous trace, inferred faults). C Geographic and geologic context of the Orce-Venta Micena sector in the Baza Basin. 1- Dolomites and limestones of the Subbetic domain (External Zones); 2- Red alluvial conglomerates; 3- Yellow lacustrine marls; 4- Lacustrine limestones; 5- Lacustrine marls, sands and gypsum; 6- Red and grey fluvial and lacustrine sands and clays; 7- Lacustrine marls and limestones; 8- Alluvial conglomerates; 9- Alluvial-deltaic conglomerates and sands, and lacustrine marls; 10- Glacis surface; 11- Recent fluvial gravels.
The archaeological sites of BLD and FN3 preserve huge lithic assemblages of Oldowan (i.e., Mode 1) tradition (Barsky et al. 2015.1, 2016.1, 2022.1; Martínez-Navarro et al. 1997; Turq et al. 1996; Toro-Moyano et al. 2011.2, 2013.1; Titton et al. 2018.1, 2021) and abundant skeletal remains of vertebrates, particularly large mammals. Some of these fossils show anthropogenic marks related to defleshing, butchering and marrow processing (Espigares et al. 2013, 2019, 2023; Palmqvist et al. 2023.1a, 2023.2b; Yravedra et al. 2021.1, 2022.2). Moreover, a deciduous tooth of Homo sp. has been unearthed from BLD, which is by the moment the earliest hominin fossil from Western Europe (Toro-Moyano et al. 2013.1).
There are several age estimates for BLD and FN3. For example, a combination of biostratigraphy, magnetostratigraphy and the U-series/ESR dating method applied to optically bleached quartz grains and fossil mammalian teeth provided age estimates of 1.43 ± 0.38 Ma for BLD and 1.19 ± 0.21 for FN3 (Duval et al. 2012; Toro-Moyano et al. 2013.1). In addition, an age of 1.50 ± 0.31 Ma was derived for FN3 based on cosmogenic nuclides (Álvarez et al. 2015.2). Moreover, the absence of suids from the rich faunal assemblages of BLD and FN3 contributes significant biochronological information (Martínez-Navarro et al., 2015.3): Suids are not present in Europe during the 1.8–1.2 Ma interval until the arrival of a derived form of Sus strozzi during the Epivillafranchian (Cherin et al. 2018.2, 2020). This species is first recorded in association with hominin remains at unit TE9 of Sima del Elefante in the Atapuerca karstic complex, which is dated to 1.22 ± 0.16 Ma on the basis of cosmogenic nuclides (Carbonell et al. 2008), and later in other sites of Jaramillo age like Untermassfeld in Germany, Vallonnet in France, or Vallparadís in Spain (Cherin et al. 2018.2, 2020; Madurell-Malapeira et al. 2010.1, 2014.2; Moullé et al. 2006). This proposal, however, has recently been challenged by Iannucci (2024) based on a distal fragment of a left suid Mt IV from a fossil collection of the XIX century. The specimen, which is housed in the Natural History Museum, London (NHMUK PV OR 27621), comes from Peyrolles, a locality of uncertain age of the French Central Massif. Although a fossiliferous level of Peyrolles has been dated by the 40Ar/39Ar method to 1.47 Ma (Nomade et al. 2014.3), it has been suggested that the short faunal list from this site is biochronologically coeval with the Upper Valdarno (Valli et al. 2006.1), which points to an age close to 1.8 Ma. Therefore, according to the evidence discussed above, the most parsimonious age for the two Orce sites, BLD and FN3, would be ~ 1.4 Ma (see review in Palmqvist et al. 2016).
The lithic assemblages unearthed from BLD and FN3 represent the whole reduction sequence. They are composed of abundant small flakes, debris, cobbles (one-third with percussion marks), cores, spheroids, and debitage made of flint, limestones and calcarenites from the site surroundings. Flint was largely exploited for flake production while limestone, although also used for flake production, was mostly employed as percussion tools (Barsky et al. 2015.1, 2016.1, 2022.1; Martínez-Navarro et al. 1997; Oms et al., 2000; Palmqvist et al., 2005; Toro-Moyano et al. 2011.2, 2013.1; Titton et al. 2018.1, 2021; Tixier et al. 1995; Turq et al. 1996). A small proportion of bones (<2%) show cut marks and percussion marks resulting from defleshing and bone fracturing for accessing their marrow contents, respectively (Espigares 2010.2; Espigares et al. 2013, 2019; Palmqvist et al. 2023.1a; Yravedra et al. 2021.1). Incisions predominate among cut marks, although scrapes, sawing marks and chop marks are also documented. These marks evidence patterns of skinning, disarticulation, defleshing, evisceration, and periosteum removal. They concentrate on the bones of large and very large ungulates, like horses (Equus altidens and Equus suessenbornensis), megacerine deer (Praemegaceros cf. verticornis), bison (Bison sp.), rhino (Stephanorhinus hundsheimensis), hippo (Hippopotamus antiquus), and elephant (Mammuthus meridionalis). Evidence of intentional bone breakage includes percussion marks, pits, notches, impact flakes, and negative flake scars generated by a direct contact of the bones with the hammerstone (Espigares et al., 2019; Yravedra et al., 2021.1).
The faunal assemblages of BLD and FN3 also preserve bones with carnivore modifications. These bones are recorded in low proportions, like those with anthropogenic marks, and include tooth marks similar in morphology, dimensions and anatomical position to those found at the late Early Pleistocene site of Venta Micena (VM) in Orce, in which tooth marks are more abundantly represented (Courtenay et al., 2023.3; Espigares, 2010.2; Espigares et al., 2019; Palmqvist et al., 2023.2b; Yravedra et al., 2021.1). The VM site, which is dated by biostratigraphy to 1.6–1.5 Ma (Martínez-Navarro, 1991; Palmqvist et al., 2022.2b), has been interpreted as a breeding den of the giant, short-faced hyena Pachycrocuta brevirostris (Arribas & Palmqvist, 1998; Espigares, 2010.2; Palmqvist & Arribas, 2001; Palmqvist et al. 1996.1, 2011.3, 2022.2b). For this reason, most tooth-marked bones of BLD and FN3 were probably also consumed by P. brevirostris (Espigares et al. 2013, 2019, 2023; Palmqvist et al., 2023.1a, 2023.2b). However, some activity of small carnivorans has also been identified in these sites (Espigares, 2010.2; Espigares et al., 2013, 2019), probably the medium-sized canid Canis mosbachensis (Courtenay et al., 2023.3; Palmqvist et al., 2023.2b). Moreover, a bone shaft from FN3 was apparently gnawed by a porcupine (Espigares et al., 2019).
The activity of P. brevirostris is particularly evident in the case of the upper archaeological level of FN3, in which hyena coprolites are abundantly preserved (Espigares, 2010.2; Espigares et al., 2019, 2023), as explained below. Apart from this large hyaenid, carnivores are well represented in BLD and FN3, including two mustelids (Martellictis ardea and Meles meles), three canids (Vulpes alopecoides, C. mosbachensis, and Lycaon lycaonoides), one ursid (Ursus etruscus), and three felids (Lynx pardinus, Megantereon whitei and Homotherium latidens) (Martínez-Navarro et al., 2010.3; Palmqvist et al., 2023.1a; Rodríguez-Gómez et al., 2016.2a, 2016.3b; Ros-Montoya et al., 2021.2). This rich carnivore guild resembles those found in other late Early Pleistocene archaeological and paleontological sites of Eurasia, such as Dmanisi in Georgia, dated to ~ 1.8 Ma (Medin et al., 2019.1; Bartolini-Lucenti et al., 2021.3, 2022.3), Pirro Nord in Italy, dated to 1.6–1.3 Ma (Petrucci et al., 2013.2), and Vallonnet in France, dated to ~ 1.1 Ma (Moullè et al., 2006), or at the early Acheulian site of ‘Ubeidiya in Israel, dated to 1.6–1.2 Ma (Martínez-Navarro et al., 2009).
The lithic assemblages of BLD and FN3, together with the inferences on the availability of animal resources and the intensity of competition among carnivores and hominins for these resources (Rodríguez-Gómez et al., 2016.2a, 2016.3b, 2022.4, 2024.1), provide interesting clues on the subsistence strategies of the first human populations that colonized Western Europe during late Early Pleistocene times (Espigares et al., 2013, 2019; Palmqvist et al., 2022a, 2023.1a; Rodríguez-Gómez et al., 2016.2a, 2017.1, 2022.4, 2024.1; Toro-Moyano et al., 2013.1).
1.1 The late Early Pleistocene site of FN3
The stratigraphy of FN3 consists of a ~ 5 m thick section formed by the alternation of sub-horizontal limestones with lutites and fine sands. This stratigraphic section was first described by Turq et al. (1996) and later by Oms et al. (2011.4), who described twelve layers (see Fig. 2A–C), numbered from bottom to top and grouped into three sedimentary cycles. Each cycle begins in a lower part constituted by greenish marly lutites and clays, as well as sands with iron oxides and carbonate nodules, and ends in an upper part characterized by a limestone/marly limestone stratum with evidence of microkarstification, together with intraclasts and diagenetic markers (Fig. 2D). These lutitic-carbonate sediments were deposited in a shallow lacustrine-to-swampy environment of variable depth, with occasional fluvial inputs (Turq et al., 1996; Oms et al., 2011.4; Espigares et al., 2023). Specifically, the clays and marly lutites were deposited under a semi-permanent water table while the sands correspond to lake shore deposits.The first sedimentary cycle is only represented by its upper part (layer 1), a whitish limestone stratum with a soft-sediment deformation structure that shows a rounded, pillow-shaped surface (Fig. 3). The second cycle (layers 2–4) and the third (layers 5–12) are both preserved complete. Layer 2 is an irregular bed of green clays that conforms to the surface of layer 1, showing a similar soft-sediment deformation structure. This layer is of variable thickness (10–20 cm) and includes small intercalations of sand. Layer 3 was deposited in concordance with layer 2 and consists of brown-whitish clays with carbonate nodules. Layers 1–3 show plastic deformation and small synsedimentary normal faults with a vertical extent of a few centimeters (Fig. 2C). In all probability, these structures were formed by earthquakes (Oms et al., 2011.4), which are frequently recorded in the Baza Basin (Alfaro et al., 1997.1, 2010.4; Gibert et al., 2005.1). Layer 4 is a whitish limestone that seems to have been less affected by deformation. The bottom of this layer conforms to the paleosurface, while its top is eroded by the deposition of the overlying sediments. The third sedimentary cycle consists of layers 5–12 (Fig. 2B). Layer 5 is of variable thickness (20–30 cm, although it can occasionally be thicker) and consists of greenish sands with lenticular shapes, which can be interpreted as the result of low-energy fluvial flows with sufficient energy to erode the unconsolidated limestones of layer 4. A high percentage (68.2%, 5466 out of 8014) of the fossil remains of large mammals unearthed from FN3 are from layer 5 (Espigares et al., 2023: Table 1). Layers 6–10 are composed of marly lutites and clays, which show differences in color and thickness. The boundary between layers 7 and 8 shows an important irregularity, which may be indicative of an erosional process resulting from subaerial emersion. Finally, layers 11–12 are horizontal whitish limestones that close the third sedimentary cycle and the stratigraphic section of FN3.Fig. 2 A Stratigraphic scheme of the layers of the Fuente Nueva-3 (FN3) site (modified from Oms et al., 2011.4; Fig. 6; Espigares et al., 2023: Fig. 2). This stratigraphy shows a succession of layers constituted by whitish nodular limestones (layers 1, 4, 11, and 12) and irregular layers of yellowish marly lutites (layers 2–3 and 6–10), as well as dark greenish sands that appear in an intermediate position (layer 5). All these layers show evidence of subaerial pedogenesis and can be grouped into three sedimentary cycles (Espigares et al., 2023; Oms et al., 2011.4; Palmqvist et al., 2023.1a). B Image showing the morphology of the upper layers (4–7), which constitute the upper archaeological level (UAL). C Image showing the morphology of the lower layers (1–3), which represent the lower archaeological level (LAL). Red lines indicate faulting. D Stratigraphic series of the FN3 siteFrom an archaeological point of view, the fertile strata of FN3 have a thickness of ~ 1.5–2 m and comprise the first seven layers, which have been grouped in two main archaeological levels (Turq et al., 1996; Martínez-Navarro et al., 1997; Oms et al., 2011.4; Espigares et al. 2013, 2019, 2023; Reinoso et al., 2020; Palmqvist et al., 2023.1a): The lower archaeological level (LAL, layers 1–3) and the upper archaeological level (UAL, layers 4–7). Fossil bones, hyena coprolites, lithic tools, and manuports have been recovered from six different layers of FN3, although most of them come from layers 2–3 of the LAL and layer 5 of the UAL (Espigares et al., 2023: Table 1). Up to the 2015 excavation season, 1367 Mode 1 lithic artifacts knapped in situ were unearthed from FN3. In addition, there are 375 unmodified stones or manuports (i.e., non-flaked, allochthonous blocks and cobbles from the Jurassic carbonate rocks of the site surroundings). Nearly half of these limestone blocks display percussion marks (Barsky et al., 2015.1, 2016.1, 2022.1), which suggests they were probably intentionally brought to the FN3 site by the hominins, who used them as percussion tools for fracturing the bones of large mammals.Fig. 3
A Stratigraphic scheme of the layers of the Fuente Nueva-3 (FN3) site (modified from Oms et al., 2011.4; Fig. 6; Espigares et al., 2023: Fig. 2). This stratigraphy shows a succession of layers constituted by whitish nodular limestones (layers 1, 4, 11, and 12) and irregular layers of yellowish marly lutites (layers 2–3 and 6–10), as well as dark greenish sands that appear in an intermediate position (layer 5). All these layers show evidence of subaerial pedogenesis and can be grouped into three sedimentary cycles (Espigares et al., 2023; Oms et al., 2011.4; Palmqvist et al., 2023.1a). B Image showing the morphology of the upper layers (4–7), which constitute the upper archaeological level (UAL). C Image showing the morphology of the lower layers (1–3), which represent the lower archaeological level (LAL). Red lines indicate faulting. D Stratigraphic series of the FN3 siteFrom an archaeological point of view, the fertile strata of FN3 have a thickness of ~ 1.5–2 m and comprise the first seven layers, which have been grouped in two main archaeological levels (Turq et al., 1996; Martínez-Navarro et al., 1997; Oms et al., 2011.4; Espigares et al. 2013, 2019, 2023; Reinoso et al., 2020; Palmqvist et al., 2023.1a): The lower archaeological level (LAL, layers 1–3) and the upper archaeological level (UAL, layers 4–7). Fossil bones, hyena coprolites, lithic tools, and manuports have been recovered from six different layers of FN3, although most of them come from layers 2–3 of the LAL and layer 5 of the UAL (Espigares et al., 2023: Table 1). Up to the 2015 excavation season, 1367 Mode 1 lithic artifacts knapped in situ were unearthed from FN3. In addition, there are 375 unmodified stones or manuports (i.e., non-flaked, allochthonous blocks and cobbles from the Jurassic carbonate rocks of the site surroundings). Nearly half of these limestone blocks display percussion marks (Barsky et al., 2015.1, 2016.1, 2022.1), which suggests they were probably intentionally brought to the FN3 site by the hominins, who used them as percussion tools for fracturing the bones of large mammals.Fig. 3 Basal limestones (layer 1) of the Fuente Nueva-3 (FN3) site showing dome-shaped deformation structures, which were probably caused by an earthquake (photographs taken during the excavation season of the year 2003). A View of the dome structure with an approximately E-W orientation. The red dashed line to the northern wall shows that the overlying layers adapt to the morphology of the limestones. B Top view of the basal limestones showing two dome-shaped structures and abundant fractures. Similar dome-shaped structures figured by Gibert et al. (2005.1) were interpreted as paleoseismites.
Basal limestones (layer 1) of the Fuente Nueva-3 (FN3) site showing dome-shaped deformation structures, which were probably caused by an earthquake (photographs taken during the excavation season of the year 2003). A View of the dome structure with an approximately E-W orientation. The red dashed line to the northern wall shows that the overlying layers adapt to the morphology of the limestones. B Top view of the basal limestones showing two dome-shaped structures and abundant fractures. Similar dome-shaped structures figured by Gibert et al. (2005.1) were interpreted as paleoseismites.
In a recent study, Espigares et al., (2023: Table 1) showed that fossil bones and lithic tools are abundantly preserved in both archaeological levels of FN3 (see Fig. 4): 2234 bones and 507 lithic tools come from the LAL, while 5780 bones and 482 tools are from the UAL. However, there are differences between both archaeological levels. For example, the density of manuports is higher in the LAL than in the UAL: 329 out of 375 manuports (87.7%) come from layers 2–3 of the LAL and only 46 (12.3%) from layer 5 of the UAL. This suggests that hominin activity concentrated in the LAL. In contrast, hyena coprolites are more abundantly represented in the UAL than in the LAL (Fig. 4): 98.2% (216 out of 220) coprolites preserved in FN3 come from layer 5 of the UAL and only three were recovered from layers 2–3 of the LAL (Espigares et al., 2023: Table 1). This suggests that hyaenid activity was particularly intense at the UAL.According to Espigares et al. (2023), the lithological differences between the LAL and UAL could tentatively explain the differences in the composition of their bone assemblages of large mammals. Specifically, the fossiliferous layers of the UAL (layers 5–7; see Fig. 2) are composed of greenish fine sands and marly mudstones (layer 5), dark brown clays (layer 6), and greenish-brown dark marly mudstones (layer 7). In contrast, the fertile layers of the LAL (layers 2–3) are composed of green clays (layer 2) and brown whitish clays with nodules (layer 3). The composition of layer 5, which preserves most of the skeletal remains from the UAL, led Espigares et al. (2023) to propose that this layer could have behaved as quicksand that became unstable when it was forced to move by the pressure exerted by very large sized animals like megaherbivores, which remains are particularly abundant in this layer. For this reason, Espigares et al. (2023) interpreted layer 5 of the UAL, which preserves a high number of hyena coprolites, as a latrine of the giant hyena P. brevirostris developed in an open-air setting.Fig. 4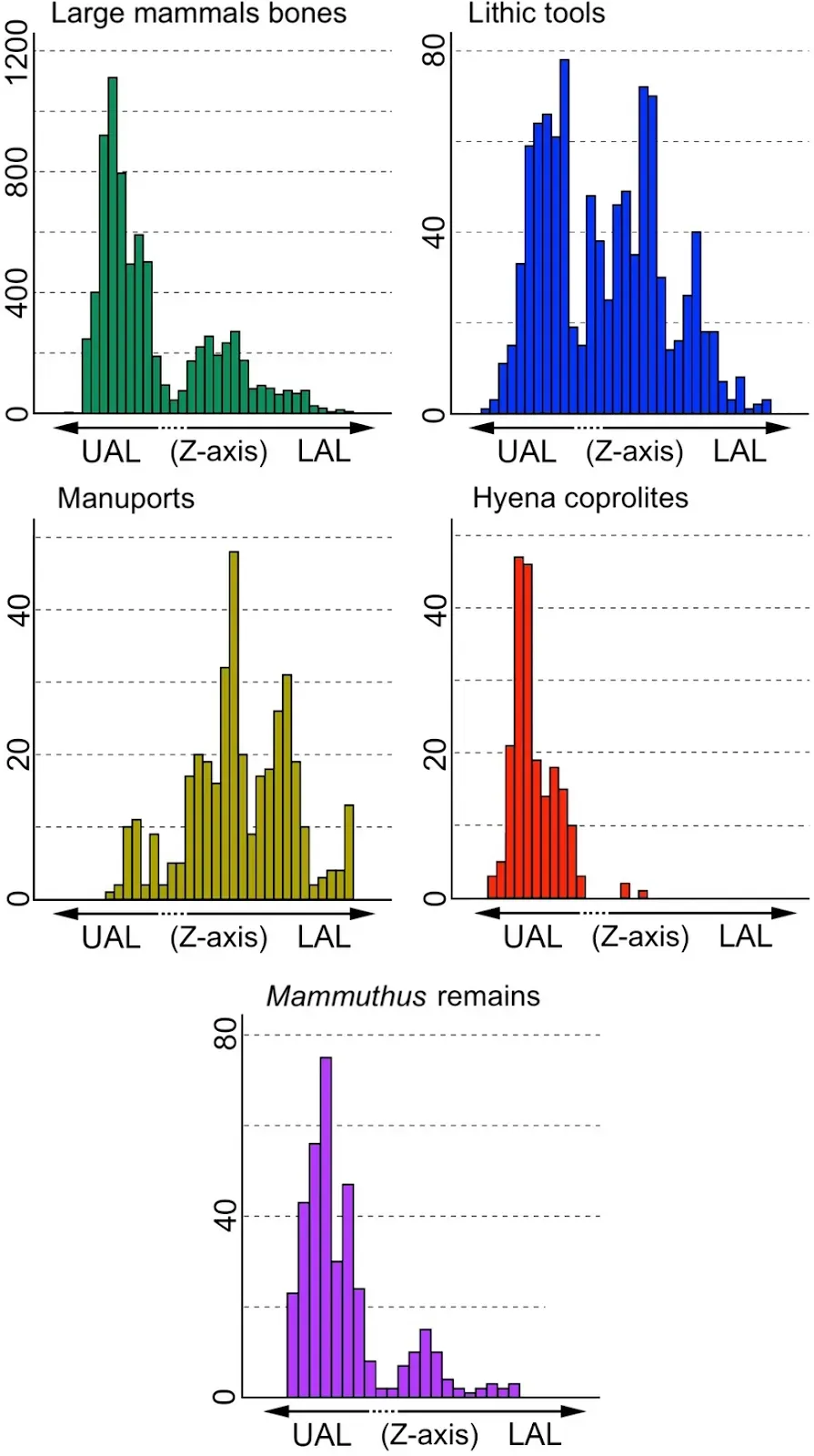 Abundance (Z-axis) in the two archaeological levels of the late Early Pleistocene site of Fuente Nueva-3 (FN3) of fossil remains (mostly large mammals; see Table 1), lithic tools (flakes, debris, cobbles, and debitage), manuports (unmodified limestone blocks intentionally brought to the site by the hominins), hyena coprolites, and skeletal remains of elephant Mammuthus meridionalis (data from Espigares et al., 2023: Figure 12 and Table 1). The limit of the layers included in the upper archaeological level (UAL, layers 4–7) and lower archaeological level (LAL, layers 2–3) is not depicted in this figure, because these layers show irregular bedding surfaces (see Fig. 2)
Abundance (Z-axis) in the two archaeological levels of the late Early Pleistocene site of Fuente Nueva-3 (FN3) of fossil remains (mostly large mammals; see Table 1), lithic tools (flakes, debris, cobbles, and debitage), manuports (unmodified limestone blocks intentionally brought to the site by the hominins), hyena coprolites, and skeletal remains of elephant Mammuthus meridionalis (data from Espigares et al., 2023: Figure 12 and Table 1). The limit of the layers included in the upper archaeological level (UAL, layers 4–7) and lower archaeological level (LAL, layers 2–3) is not depicted in this figure, because these layers show irregular bedding surfaces (see Fig. 2)
1.2 Quicksand as a potential deadly trap for megafauna
Quicksand can potentially be a deadly trap for wildlife. Given its composition (fine sand, silt and salt water), quicksand is extremely sensitive to even small changes in stress, which can result in significant variations in its steady-state viscosity, thus causing the sediment to liquefy. For this reason, the higher the stress, the more fluid the quicksand becomes, so movement by a trapped body causes it to sink (Khaldoun et al., 2005.2, 2006.2). After an animal sinks into quicksand, the viscosity of the sediment increases due to the separation of the quicksand into two phases: a water-rich phase and a sand-rich phase (Khaldoun et al., 2005.2). This causes an animal trapped in quicksand to sink deeper when it moves trying to escape from the quicksand trap (Khaldoun et al., 2005.2). In such an environment, viscosity can reach such high levels that an animal may require a force up to three times its weight to free itself from the sediment (Kadau et al., 2009.1).
If artesian conditions exist, quicksand is formed by spring water issuing from an underlying permeable stratum fed by a head of water at higher elevation (Matthes, 1953). Surface quicksand is likely to occur in hilly country, especially in regions of calcareous rocks like the Baza Basin, which often contain solution openings, thus favoring the occurrence of springs (Patchick, 1966). In these environments, the sun bakes a thin crust on the sand (Matthes, 1953), which hides the risk of this substrate for animals moving in the vicinity.
The coefficient of friction and the penetration hardness of a quicksand depend on the water contents it incorporates (Fall et al., 2014.4; Liefferink et al., 2020.2). At low water contents the sand shows a pendular regime and can withstand relatively high loads before reaching the critical penetration hardness. Deformation of the granular material causes a reorientation of the grains, including the breaking and rebuilding of capillary liquid bonds. An increase in water content causes coalescence of the liquid bonds and a shift to a capillary regime, which further decreases the plastic response of the granular material. As a result, the coefficient of friction increases, resulting in an increased risk of entrapment for heavy animals with large limb sections. In addition, the high body mass of a trapped megaherbivore like an elephant makes it more difficult for it to counteract the negative suction of the quicksand to lift its pillar-like legs out of the sand (see Ren et al., 2010.5). In this scenario, an increase in salinity increases the cohesion and viscosity between the sand grains, which in turn increases the hardness of penetration into the quicksand (Fall et al., 2014.4). In a warm region like the Baza Basin, the evaporation of water from a sand with salt water results in the formation of salt crystals that fill the spaces between the sand grains. This process binds the sand grains together more tightly, creating a stiffer material that is more resistant to penetration stresses. The increase in strength is particularly noticeable in the pendular regime, where the amount of water (and thus dissolved salt) is relatively low but sufficient to form liquid bridges which, when evaporating, leave salt crystals that function as solid bridges in the surface crust. This is a double-edged trap because the surface crust is fragile. For this reason, if the fracture limit at the surface is exceeded (Nguyen et al., 2016.4), a death trap occurs when the limbs of a large sized animal move into the sand layer with an elevated water content, where the penetration hardness is lower and the friction is higher, thus trapping the animal. This would explain the difference of behavior between the LAL and UAL of FN3, and also why hyenas and hominins were able to roam the surface of quicksand without facing excessive risk of sinking into it, as explained below.
Experimental data on quicksand from salt-lake environments composed of fine sand, clay and salt water, similar in composition to those found in the sediments deposited in the oligosaline lacustrine systems recorded at the Orce sites (García-Aguilar et al., 2014, 2015; Palmqvist et al., 2022a), show that the quicksand acts as a trap for the animals caught. Espigares et al. (2023) proposed that this scenario would explain the high abundance of skeletal remains of megaherbivores, particularly elephants, in the UAL of FN3, and also the high density of hyena coprolites, because the half-sunken carcasses of the animals that regularly got stuck in the muddy sands would have attracted the scavenging hyenas.
Another way of fluidization for a quicksand can be vibrations from an earthquake (Kadau et al., 2009.1). Of interest here, paleo-seismites are frequently recorded in the Orce sediments and throughout the whole Baza Basin, which shows that this region was tectonically and seismically active during the Quaternary (Alfaro et al., 1997.1, 2010; García-Aguilar et al., 2014; Gibert et al., 2005.1). In fact, the dome-shaped surface of the limestone layers of FN3 can be interpreted as resulting from earthquakes (see Fig. 3).
Most research on quicksand focuses on the danger it poses to man-made structures (Craig, 1997.2; Elbaz et al., 2016.5; Sun, 2016.6; Xu et al., 2009.2). However, with the only exception of Hungerbühler (1998.1), who proposed a quicksand-like trap as an explanation for the finding of 17 partial skeletons of the Late Triassic prosauropod dinosaur Sellosaurus gracilis, as far as we know no paleo-quicksand has been reliably identified in the sedimentological record. For this reason, it is of great interest to test if layer 5 of the UAL of FN3 could have behaved as a quicksand in which very large animals became entrapped, as proposed by Espigares et al. (2023).
1.3 Objectives
The two main goals of this study are: (i) to test for statistical differences in the composition of the fossil assemblages of large mammals preserved in the LAL and UAL of FN3; and (ii) to analyze particle size in the main fertile layers of the LAL (layers 2–3) and UAL (layer 5), which preserve most skeletal remains of large mammals unearthed from the FN3 site, in order to test the hypothesis of entrapment of megaherbivores in quicksand proposed by Espigares et al. (2023) for layer 5 of the UAL.
Palmqvist, P., Campaña, I., Granados, A. et al.
The late Early Pleistocene site of Fuente Nueva-3 (Guadix-Baza Depression, SE Spain): a hyena latrine developed on a quicksand trap for megaherbivores? J Iber Geol (2024). https://doi.org/10.1007/s41513-024-00241-1
Copyright: © 2024 The authors.
Published by Springer Nature Ltd. Open access.
Reprinted under a Creative Commons Attribution 4.0 International license (CC BY 4.0)



Genesis is best regarded as an illustration of what our primitive ancestors once believed before we knew any better, and how ignorant guesses and superstitions are no match for science in gaining an understanding of the world.
What Makes You So Special? From The Big Bang To You

How did you come to be here, now? This books takes you from the Big Bang to the evolution of modern humans and the history of human cultures, showing that science is an adventure of discovery and a source of limitless wonder, giving us richer and more rewarding appreciation of the phenomenal privilege of merely being alive and able to begin to understand it all.
Available in Hardcover, Paperback or ebook for Kindle
Ten Reasons To Lose Faith: And Why You Are Better Off Without It

This book explains why faith is a fallacy and serves no useful purpose other than providing an excuse for pretending to know things that are unknown. It also explains how losing faith liberates former sufferers from fear, delusion and the control of others, freeing them to see the world in a different light, to recognise the injustices that religions cause and to accept people for who they are, not which group they happened to be born in. A society based on atheist, Humanist principles would be a less divided, more inclusive, more peaceful society and one more appreciative of the one opportunity that life gives us to enjoy and wonder at the world we live in.
Available in Hardcover, Paperback or ebook for Kindle

No comments :
Post a Comment
Obscene, threatening or obnoxious messages, preaching, abuse and spam will be removed, as will anything by known Internet trolls and stalkers, by known sock-puppet accounts and anything not connected with the post,
A claim made without evidence can be dismissed without evidence. Remember: your opinion is not an established fact unless corroborated.