

The problem with trying to cling to the daft notion that Earth is only 10,000 years old and that Homo sapiens didn't evolve but were magicked out of dirt without ancestors or biological relatives, is that science keeps on proving that idea wrong, by discovering other hominins and a history that occurred in that 99.9975% of Earth's history that happened before 10,000 years ago.
So, here is some more evidence for creationists to ignore, lie about or misrepresent. It concerns a population of archaic Eurasian hominin know to science as the Denisovans, which are yet to be given a definitive species name because too few remains have been discovered, other than their DNA.
Their DNA has been isolated from a finger bone discovered in the Denisova Cave in the Altai mountains of Siberia and it shows that they interbred both with Neanderthals and with early Homo sapiens as the latter spread across Asia into their range.
Tell me all about the Denisovans, their origins and distribution, and the evidence of interbreeding with Neanderthals and early Homo sapiens. Denisovans: Origins and DistributionAnd now, a team of palaeoanthropologists have found evidence that they survived on the Tibetan plateau for about 160,000 years. The research team was led by Lanzhou University, China, the University of Copenhagen, Denmark, the Institute of Tibetan Plateau Research, CAS, China, and involved the University of Reading. They studied more than 2,500 bones from the Baishiya Karst Cave on the high-altitude Tibetan Plateau, one of the only two places where Denisovans are known to have lived.
Origins:
Denisovans are an extinct group of archaic humans closely related to Neanderthals. They were identified from genetic evidence rather than extensive fossil records. The first evidence of Denisovans came from a finger bone fragment discovered in the Denisova Cave in the Altai Mountains of Siberia in 2008. This cave has provided a treasure trove of information about this mysterious group.
Distribution:
Denisovans are believed to have lived in a wide geographical area across Asia. Their remains have been primarily found in Denisova Cave, but genetic evidence suggests they were more widespread. Denisovan DNA is found in varying degrees in the genomes of present-day populations in Oceania, Southeast Asia, and East Asia, indicating a broad range.
Fossil Evidence
The fossil record for Denisovans is sparse. Key discoveries include:
- Denisova Cave, Siberia: The primary site where Denisovan fossils have been found. These include a finger bone, teeth, and a fragment of a long bone.
- Baishiya Karst Cave, Tibet: A jawbone with Denisovan-like features was discovered here, indicating Denisovans lived at high altitudes.
Genetic Evidence and Interbreeding
Genetic Studies:
Denisovans were identified through genetic analysis of mitochondrial DNA (mtDNA) and nuclear DNA extracted from the finger bone found in Denisova Cave. This analysis revealed that Denisovans were a distinct group, sharing a common ancestor with Neanderthals and modern humans. Interbreeding with Neanderthals:
Genetic evidence shows that Denisovans and Neanderthals interbred. Studies indicate that:
Interbreeding with Early Homo sapiens:
- Neanderthals contributed to the Denisovan gene pool.
- Denisovans and Neanderthals shared a common ancestor around 600,000 years ago.
- There were at least two episodes of interbreeding between Denisovans and Neanderthals.
Evidence of Denisovan admixture in modern human populations suggests several interbreeding events:
- Oceania: Modern Melanesians and Aboriginal Australians have the highest Denisovan ancestry, with up to 5% of their DNA derived from Denisovans.
- East Asia and Southeast Asia: Populations in these regions also carry Denisovan genetic markers, though in smaller percentages.
- Tibetan Plateau: The EPAS1 gene variant, which helps with high-altitude adaptation in Tibetans, is inherited from Denisovans.
Impact on Modern Humans
Denisovan genes have influenced modern human biology in various ways:
- Immune System: Some Denisovan genes may have helped early humans adapt to new pathogens.
- Altitude Adaptation: The EPAS1 gene variant helps Tibetans survive at high altitudes with low oxygen levels.
- Physical Traits: Some genetic contributions from Denisovans might affect physical traits, though these effects are still being studied.
Conclusion
The Denisovans were a significant branch of the human family tree, contributing to the genetic diversity of modern humans through interbreeding. Despite the limited fossil record, genetic analysis has provided substantial insight into their interactions with Neanderthals and early Homo sapiens, and their legacy lives on in the DNA of many contemporary human populations.
Why have they not been given a scientific binomial?
Denisovans have not been given a scientific binomial (i.e., a formal species name) primarily due to the limited fossil evidence and ongoing debates within the scientific community regarding their classification. Here are some of the main reasons:
Limited Fossil Evidence
The fossil record for Denisovans is extremely sparse. So far, the main Denisovan fossils include:
These remains are not enough to comprehensively understand their physical morphology and make definitive comparisons with other human species. Without more complete fossil remains, it is difficult to describe Denisovans as a distinct species.
- A finger bone
- Several teeth
- A jawbone fragment
Genetic Evidence Over Morphology
Denisovans were primarily identified through genetic analysis rather than through traditional paleontological methods. This reliance on DNA rather than a comprehensive set of physical characteristics complicates the process of giving them a formal species name. Traditional taxonomy relies heavily on physical morphology, which is lacking in the case of Denisovans.
Ongoing Debate and Complexity
There is still considerable debate among scientists about how to classify Denisovans. Some researchers propose that Denisovans, Neanderthals, and modern humans are subspecies of Homo sapiens, given their ability to interbreed and produce fertile offspring. Others argue that Denisovans should be classified as a separate species due to their genetic distinctiveness.
Interbreeding and Gene Flow
The evidence of interbreeding between Denisovans, Neanderthals, and early modern humans adds complexity to their classification. The gene flow between these groups blurs the lines between distinct species and subspecies. This complexity makes it challenging to assign a clear binomial name.
Precedent and Caution
Naming a new species is a significant step that requires a high degree of certainty. Scientists often proceed with caution, especially when the evidence is limited or still emerging. Given that Denisovans have only been known for a relatively short time (since 2010), the scientific community may be waiting for more evidence before making a formal decision.
Conclusion
The absence of a scientific binomial for Denisovans reflects the complexities and uncertainties in classifying this group based on the currently available evidence. As more fossils are discovered and our understanding of Denisovan genetics and morphology improves, it may become possible to formally name and classify them within the broader human lineage.
The evidence of their existence for that long on the Tibetan Plateau is indirect - bones of animals they butchered for food, as shown by the cut marks on them have been dated that far back.
Their findings are the subject of a paper in the journal Nature and a new release from the University of Reading, Berkshire, UK:
Extinct humans survived on Tibetan plateau for 160,000 years
Bone remains found in a Tibetan cave 3,280 m above sea level indicate an ancient group of humans survived here for many millennia, according to a new study published in Nature.
The Denisovans are an extinct species of ancient human that lived at the same time and in the same places as Neanderthals and Homo sapiens. Only a handful Denisovan remains have ever been discovered by archaeologists. Little is known about the group, including when they became extinct, but evidence exists to suggest they interbred with both Neanderthals and Homo sapiens.
A research team led by Lanzhou University, China, the University of Copenhagen, Denmark, the Institute of Tibetan Plateau Research, CAS, China, and involving the University of Reading studied more than 2,500 bones from the Baishiya Karst Cave on the high-altitude Tibetan Plateau, one of the only two places where Denisovans are known to have lived.
Their new analysis, published today (Wednesday, 3 July) in Nature, has identified a new Denisovan fossil and shed light on the species’ ability to survive in fluctuating climatic conditions — including the ice age — on the Tibetan plateau from around 200,000 to 40,000 years ago.
We were able to identify that Denisovans hunted, butchered and ate a range of animal species. Our study reveals new information about the behaviour and adaptation of Denisovans both to high altitude conditions and shifting climates. We are only just beginning to understand the behaviour of this extraordinary human species.
Dr Geoff Smith, co-author
Department of Archaeology
University of Reading, Reading, UK.
Dietary diversity
Bone remains from Baishya Karst Cave were broken into numerous fragments preventing identification. The team used a novel scientific method that exploits differences in bone collagen between animals to determine which species the bone remains came from.
Zooarchaeology by Mass Spectrometry (ZooMS) allows us to extract valuable information from often overlooked bone fragments, providing deeper insight into human activities.
Dr Huan Xia, co-lead author
College of Earth and Environmental Sciences
Lanzhou University, Lanzhou, China.
The research team determined that most of the bones were from blue sheep, known as the bharal, as well as wild yaks, equids, the extinct woolly rhino, and the spotted hyena. The researchers also identified bone fragments from small mammals, such as marmots, and birds.
Current evidence suggests that it was Denisovans, not any other human groups, who occupied the cave and made efficient use of all the animal resources available to them throughout their occupation.
Dr Jian Wang, of Lanzhou University, co-lead author.
College of Earth and Environmental Sciences
Lanzhou University, Lanzhou, China.
Detailed analysis of the fragmented bone surfaces shows the Denisovans removed meat and bone marrow from the bones, but also indicate that humans used them as raw material to produce tools.
A new Denisovan fossil
The scientists also identified one rib bone as belonging to a new Denisovan individual. The layer where the rib was found was dated to between 48,000 and 32,000 years ago, implying that this Denisovan individual lived at a time when modern humans were dispersing across the Eurasian continent. The results indicate that Denisovans lived through two cold periods, but also during a warmer interglacial period between the Middle and Late Pleistocene eras.
 Denisovan rib.Credit: Dongju Zhang’s group (Lanzhou University).
Denisovan rib.Credit: Dongju Zhang’s group (Lanzhou University).
Together, the fossil and molecular evidence indicates that Ganjia Basin, where Baishiya Karst Cave is located, provided a relatively stable environment for Denisovans, despite its high-altitude. The question now arises when and why these Denisovans on the Tibetan Plateau went extinct.
Dr Frido Welker, co-corresponding author
Globe Institute
University of Copenhagen, Copenhagen, Denmark.
Xia, H., Zhang, D., Wang, J. et al.
Middle and Late Pleistocene Denisovan subsistence at Baishiya Karst Cave.
Nature (2024). https://doi.org/10.1038/s41586-024-07612-9

AbstractIf the existence of a third contemporaneous species of Homo in Eurasia, in addition to the Neanderthals, doesn't suggest to creationists that their legendary magic creation of the species as a single couple, magically made from dirt, 10,000 years ago might possibly have a flaw, then maybe that fact that this third species, the Denisovans, were living on the Tibetan Plateau 160,000 years ago might just do the trick.
Genetic and fragmented palaeoanthropological data suggest that Denisovans were once widely distributed across eastern Eurasia1,2,3. Despite limited archaeological evidence, this indicates that Denisovans were capable of adapting to a highly diverse range of environments. Here we integrate zooarchaeological and proteomic analyses of the late Middle to Late Pleistocene faunal assemblage from Baishiya Karst Cave on the Tibetan Plateau, where a Denisovan mandible and Denisovan sedimentary mitochondrial DNA were found3,4. Using zooarchaeology by mass spectrometry, we identify a new hominin rib specimen that dates to approximately 48–32 thousand years ago (layer 3). Shotgun proteomic analysis taxonomically assigns this specimen to the Denisovan lineage, extending their presence at Baishiya Karst Cave well into the Late Pleistocene. Throughout the stratigraphic sequence, the faunal assemblage is dominated by Caprinae, together with megaherbivores, carnivores, small mammals and birds. The high proportion of anthropogenic modifications on the bone surfaces suggests that Denisovans were the primary agent of faunal accumulation. The chaîne opératoire of carcass processing indicates that animal taxa were exploited for their meat, marrow and hides, while bone was also used as raw material for the production of tools. Our results shed light on the behaviour of Denisovans and their adaptations to the diverse and fluctuating environments of the late Middle and Late Pleistocene of eastern Eurasia.
Main
Ancient DNA analysis of several hominin fossils from Denisova Cave, Russia, has revealed the existence of a sister lineage of Neanderthals in eastern Eurasia, the so-called Denisovans1. On the basis of the Denisovan genetic ancestry present in several East, South and Southeast Asian populations5,6, it is inferred that Denisovans were widespread in eastern Eurasia during the Late Pleistocene1. The Xiahe mandible (named Xiahe 1) and Denisovan sedimentary mitochondrial DNA (mtDNA) from Baishiya Karst Cave (hereafter, BKC; 3,280 metres above sea level; Extended Data Fig. 1) in Ganjia Basin on the northeastern Tibetan Plateau support this assertion3,4. Together, they show that Denisovans occupied BKC from at least 160 thousand years ago (ka) to around 60 ka, and possibly up to around 45 ka3,4.
Archaeological excavations at BKC have revealed a well-preserved stratigraphy containing a rich lithic and faunal assemblage, which provides evidence of hominin occupation from at least around 190 ka to about 30 ka (ref. 4). However, most Pleistocene sites on the Tibetan Plateau, such as the Jiangjunfu 01 site7, have yielded only a few fragmentary bone specimens. Besides the small faunal assemblage from the 151 site in the Qinghai Lake Basin, which was occupied during the last deglaciation period8, no other zooarchaeological or palaeontological data9 are available for the Middle and Late Pleistocene on the Tibetan Plateau (Supplementary Information sections 1 and 2). Therefore, BKC provides a unique opportunity to study archaic hominin subsistence strategies and the faunal ecology in which it was embedded, on the high-altitude Tibetan Plateau.
We built an extended proteomic reference database of mammalian species present in or around the Tibetan Plateau through liquid chromatography with tandem mass spectrometry (LC–MS/MS) analysis (Supplementary Information section 3 and Supplementary Data 3 and 4), to allow for subsequent high-throughput zooarchaeology by mass spectrometry (ZooMS) analysis of 1,857 bone and dental specimens from BKC (Supplementary Table 2.1). Next, we integrated taxonomic identification through ZooMS with zooarchaeological data from a larger number of faunal remains (n = 2,567; Supplementary Table 2.1 and Extended Data Fig. 2), some of which were already taxonomically identified through morphological observations. Together, this dataset provides a novel picture of the palaeoecology and the subsistence strategies of Denisovans on the Tibetan Plateau.
Composition of the faunal community at BKC
By combining morphological and ZooMS identifications, we taxonomically identify 2,005 (78.1%) of the analysed 2,567 faunal specimens (Extended Data Fig. 2, Supplementary Data 5 and Supplementary Information section 4). Our results show that caprines (Caprinae), mostly bharal (Pseudois nayaur), dominate the faunal assemblage (Extended Data Fig. 3 and Supplementary Information section 4.2). The high proportion of bovids—for example, Caprinae, wild yak (Bos cf. mutus) and Tibetan gazelle (Procapra cf. picticaudata)—and equids (Equus sp.) throughout the stratigraphy reveals a grass-dominated landscape in the Ganjia Basin during the late Middle and Late Pleistocene (Supplementary Data 5A and Supplementary Information section 4.1). The presence of forest-shrub species, such as red deer (Cervus elaphus), musk deer (Moschus sp.), groove-toothed flying squirrel (Aeretes melanopterus) and porcupine (Hystrix cf. subcristata) (Supplementary Data 5A), reflects the presence of small-scale mosaic forest-shrub habitats (Supplementary Information section 4.1), similar to the modern-day foothills and river valleys in the basin4. In addition, various carnivores (for example, spotted hyena (Crocuta crocuta ultima), wolf (Canis lupus), Tibetan fox (Vulpes ferrilata) and snow leopard (Panthera cf. uncia)) and birds (for example, golden eagle (Aquila chrysaetos) and common pheasant (Phasianus colchicus)) were also present (Supplementary Data 5A).
At present, little is known about faunal community change on the Tibetan Plateau during the Middle and Late Pleistocene (Supplementary Information section 1). Notably, we only identify extinct large carnivores (Crocuta sp.) and megaherbivores (woolly rhinoceros (Coelodonta sp.)) below layer 6 (Extended Data Fig. 3 and Supplementary Fig. 4.2). Although the sample sizes for younger layers are smaller than those for layer 10 (the modelled maximum age range is around 109 ka to more than 225 ka; Supplementary Table 2.2) and layer 11 (Extended Data Table 1), the current data suggest that there was a notable change in the composition of the faunal community around BKC during the formation of layers 6 and 5 (the modelled maximum age range is around 60–104 ka, Supplementary Table 2.2). In addition, our data also document an increase in the proportion of Caprinae over time, alongside a decrease in the proportion of Bos sp. We cannot tell whether these changes are the result of shifts in hominin foraging strategies or more specifically related to changes in the surrounding environment (Supplementary Information section 4.3). Nevertheless, the persistent presence of bovids and equids (Supplementary Fig. 4.2) combined with small variations in taxonomic diversity (Supplementary Fig. 4.3) throughout the stratigraphy suggest a generally stable open environment in Ganjia Basin.
Hominins were the primary accumulators
The surfaces of bone specimens from BKC are very well preserved, and most specimens (n = 1,616, 88.7% of n = 1,821) are within weathering stages 1 or 2, providing limited evidence for sub-aerial weathering. Traces of rodent, carnivore and anthropogenic activities were identified (Supplementary Information section 5.2 and Extended Data Fig. 4a). Rodent gnawing is limited (n = 3, 0.1%; Fig. 1a), with a slightly higher proportion of carnivore modified bones (n = 16, 0.8%; Fig. 1a), although carnivore coprolites are absent. By contrast, a larger proportion of the faunal assemblage exhibits evidence of anthropogenic modifications (n = 386, 19.3%; Fig. 1a). Most of these specimens were identified through ZooMS (Extended Data Fig. 4b). Cut marks and percussion notches were identified on both herbivore and some large carnivore (for example, Crocuta sp.) bones (Supplementary Table 5.2). The higher proportion of anthropogenically modified bones and the presence of stone artefacts in each layer4 suggests that the BKC faunal assemblage accumulated mainly through hominin activities (Supplementary Information section 5.2).
[…]Fig. 2: Examples of anthropogenically modified faunal specimens and bone tools. a, Aquila chrysaetos right humerus (layer 4) with superficial and straight cut mark clusters, associated with the removal of feathers. b, Crocuta crocuta ultima atlas (layer 10a), with an oblique cut mark generated during disarticulation. c, Marmota sp. (ZooMS taxon ID) radius diaphysis (layer 9), with a negative conchoidal medullary flake scar (black triangle) produced by anthropogenic breakage. d, A possible retoucher (layer 11). Equus sp. right lower P2 with a set of scrape marks on its buccal surface. e, Expedient bone tool (ZooMS taxon ID: Caprinae; layer 10b). This humerus diaphysis is deliberately shaped by continuous direct percussion (indicated by black triangles in the magnified image on the right) on its cortical surface. For all panels, the enlarged images (right in a,b,d,e and bottom in c) are magnifications of the regions denoted with dotted lines in the main images. Except where noted, taxonomic identifications are from morphological analysis. Scale bars, 2 cm (a–d, main images), 1 cm (e, main image) and 1 mm (all magnified images).
a, Aquila chrysaetos right humerus (layer 4) with superficial and straight cut mark clusters, associated with the removal of feathers. b, Crocuta crocuta ultima atlas (layer 10a), with an oblique cut mark generated during disarticulation. c, Marmota sp. (ZooMS taxon ID) radius diaphysis (layer 9), with a negative conchoidal medullary flake scar (black triangle) produced by anthropogenic breakage. d, A possible retoucher (layer 11). Equus sp. right lower P2 with a set of scrape marks on its buccal surface. e, Expedient bone tool (ZooMS taxon ID: Caprinae; layer 10b). This humerus diaphysis is deliberately shaped by continuous direct percussion (indicated by black triangles in the magnified image on the right) on its cortical surface. For all panels, the enlarged images (right in a,b,d,e and bottom in c) are magnifications of the regions denoted with dotted lines in the main images. Except where noted, taxonomic identifications are from morphological analysis. Scale bars, 2 cm (a–d, main images), 1 cm (e, main image) and 1 mm (all magnified images).
A new Denisovan individual
During ZooMS screening of the unidentifiable fragments, one rib specimen was identified as Homininae (Fig. 3a and Extended Data Fig. 6a). The specimen contains 14 peptide markers of collagen type I (COL1) matching Homininae, as well as one peptide marker unique to Hominoidea (Supplementary Data 4 and Extended Data Fig. 6a). Considering the current and past geographical distribution of other great apes17, in particular the genus Pan, this specimen could be confirmed as Homo sp. We therefore named this hominin specimen Xiahe 2 (field number, BSY-19-B896-1; ZooMS number, BSY-941). Xiahe 2 was broken into two pieces during excavation and belongs to the distal part of a rib (51.5 mm in length). The Xiahe 2 specimen comes from layer 3 of T3, which has been dated4 to 48–32 ka (Supplementary Table 2.2). The glutamine deamidation values of Xiahe 2 (acid COL1α1 508–519 = 0.52 and acid COL1α1 435–453 = 0.46) are similar to that of other specimens from layer 3 and specimens directly radiocarbon-dated to around 50–30 ka (Extended Data Fig. 6b). However, the deamidation values of Xiahe 2 are different from those of modern samples (Extended Data Fig. 6b), suggesting that the age of Xiahe 2 is consistent with the age of layer 3 (48–32 ka).Further shotgun proteomic analyses provide more specific information about the taxonomic attribution of Xiahe 2. We reconstructed 4,597 amino acid positions for the Xiahe 2 specimen across the 21 protein sequences used for phylogenetic analysis (14.5% of the total concatenated protein sequence alignment; Supplementary Table 4.1 and Supplementary Data 6). This is a considerably larger proteome than the six endogenous proteins that were used for the phylogenetic analysis of the Xiahe 1 mandible3. The nodes in the phylogenetic tree have high support values through both maximum likelihood and Bayesian methods, with Xiahe 2 consistently falling together with the published high-coverage Denisovan genome (Fig. 3b). As such, it can be determined that the Xiahe 2 individual is, among the available reference individuals, most closely related to the D3 Denisovan high-coverage individual. The topology and the placement of the Xiahe 2 specimen are similar to the results obtained for the previous analysis of the Xiahe 1 mandible3. The discovery of the Xiahe 2 Denisovan extends the fossil evidence for the presence of Denisovans from the late Middle Pleistocene well into the Late Pleistocene at BKC, in agreement with the Denisovan sedimentary mtDNA recovered from layer 3 at BKC4.Fig. 3: The Xiahe 2 specimen, a Homo sp. rib specimen discovered through ZooMS screening.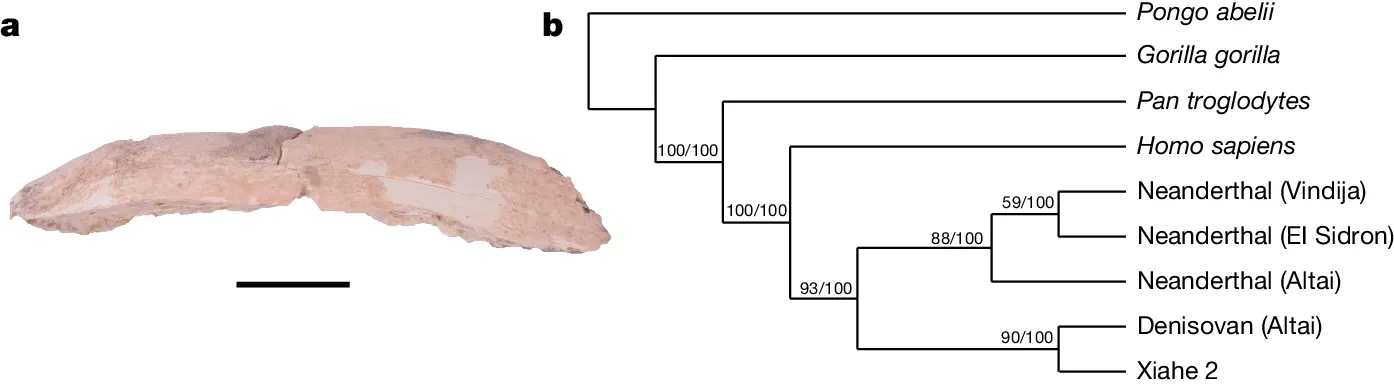 a, Photograph of the Xiahe 2 specimen. Scale bar, 1 cm. b, Phylogenetic tree for the Xiahe 2 specimen and reference proteomes. Support values at nodes are shown for the maximum likelihood and Bayesian analysis, respectively.
a, Photograph of the Xiahe 2 specimen. Scale bar, 1 cm. b, Phylogenetic tree for the Xiahe 2 specimen and reference proteomes. Support values at nodes are shown for the maximum likelihood and Bayesian analysis, respectively.
[…]
Discussion and conclusion
Previous studies show that BKC is currently the only well-preserved cave site on the Tibetan Plateau that spans the late Middle to Late Pleistocene3,4 (Supplementary Information section 1). The Xiahe 1 mandible and sedimentary mtDNA analyses reveal that Denisovans occupied the cave at least around 160 ka, 100 ka (layer 7) and 60 ka (layer 4), and possibly as late as 45 ka (end layer 4)3,4. The Xiahe 2 rib identified here, and Denisovan sedimentary mtDNA discovered from layer 3 (ref. 4), show that Denisovan occupation occurred at the site until at least 48–32 ka. Layers 10 and 11 at BKC provide the richest archaeological remains in the cave, including more than 60% of the bone specimens analysed here (Supplementary Table 2.1), but unfortunately without any hominin remains or sedimentary mtDNA to ascertain the biological identity of the occupants so far. However, the deamidation values obtained for Xiahe 1, COL1α1 508–519 (0.04) and COL1α1 435–453 (0.00) are, exclusively, within the range of those observed for bone specimens from layers 10 and 11 (Fig. 4f). In addition, Xiahe 1 has a minimum U-series age of around 160 ka (ref. 3), which also corresponds to the chronological age of layer 10 or below (Fig. 4g). Although this does not show that Xiahe 1 definitely derives from layers 10, 11 or an older layer at BKC, it does show that the Xiahe Denisovans were the most likely occupants during the formation of these layers.


More likely though, it will have creationists trawling their favourite disinformation sites looking for a way to misrepresent the science, lie about the scientists or dismiss their findings as based on falsified dating, because creationism is not an evidence-based superstition; it exists despite the evidence by selling the cult the daft notion that, although they know little or nothing of the subject, their opinions is still better than that of the experts because they are privy to a special form of knowledge called 'faith', i.e., they believe that they're told to believe and think it’s wrong to question it.
Meanwhile the whole fascinating story of their species and our relatives is closed to creationists because understanding it would be an existential threat and might them feel less important than they think they are.
What Makes You So Special? From The Big Bang To You

Ten Reasons To Lose Faith: And Why You Are Better Off Without It


No comments :
Post a Comment
Obscene, threatening or obnoxious messages, preaching, abuse and spam will be removed, as will anything by known Internet trolls and stalkers, by known sock-puppet accounts and anything not connected with the post,
A claim made without evidence can be dismissed without evidence. Remember: your opinion is not an established fact unless corroborated.