
One common way creationist apologists attempt to mislead the scientifically uninformed is by claiming that the Laws of Thermodynamics are somehow relevant to the evolution of information within a species' genome. They argue that any increase in genetic information would violate both the Second and Third Laws of Thermodynamics—asserting that increased biological complexity equates to a decrease in entropy (disorder), and that new information is akin to energy and thus cannot increase due to the Law of Conservation.
This argument is fundamentally flawed on several levels but continues to be repeated despite being repeatedly refuted by both biologists and physicists. First, it completely ignores the fact that Earth is not a closed system. The input of energy from the Sun, for example, allows local decreases in entropy (such as in the formation of complex biological structures) while the total entropy of the universe still increases, fully complying with the Second Law. The Third Law, which relates to the entropy of systems at absolute zero, is entirely irrelevant to biological evolution.
Second, the idea that genetic information is conserved like energy is a misrepresentation. Genetic information can and does change in multiple ways through mutation. A mutation can involve the loss of information (e.g. deletion of a DNA segment), a change in information (e.g. substitution of one or more nucleotides), or an increase in information (e.g. insertion of additional sequences, or the movement of transposable elements—“jumping genes”—to new locations in the genome). None of these processes require a change in the total amount of matter or energy; they simply involve the rearrangement of existing molecular components. Any local increase in biological order is offset by energy expenditure elsewhere, typically via the hydrolysis of ATP to ADP and phosphate within metabolic pathways.
Moreover, these objections rest on the false assumption that evolution is about the quantity of information. In reality, it is the function and meaning of genetic information that drives evolutionary change. A sequence of DNA that once encoded a protein with one function can, through mutation and natural selection, take on a new function entirely—a process known as exaptation.
A well-known example is the evolution of the mammalian middle ear bones. In ancestral fish, certain jawbones played a structural role in the jaw joint. Over time, in early synapsids, these bones were repurposed and miniaturised to become part of the auditory system, transmitting sound vibrations from the eardrum to the cochlea.
Other compelling examples of exaptation include:
- Crystallin proteins in the eye lens, which originally served as stress-response proteins (e.g. enzymes or heat shock proteins) before being co-opted for their optical clarity and stability in the lens.
- Feathers in birds, which almost certainly evolved initially for thermal regulation or display in theropod dinosaurs and were only later adapted for flight.
- The wings of insects, which may have evolved from gill-like structures in aquatic ancestors before becoming functional appendages for flight.
A recent discovery adds another example to this growing list. Researchers at Kobe University in Japan found that a gene currently involved in regulating root development in vascular plants is also present in liverworts, which do not possess true roots. In liverworts, the gene plays a role in organ development. This suggests that the gene existed in early land plants and was later exapted to a different developmental role in vascular plants.
The team, led by Professors Fukaki Hidehiro and Ishizaki Kimitsune, published their findings in the journal New Phytologist, and the study is further explained in a news release from Kobe University:
What information do you have on the origins and role of the RLF gene in plants? The RLF gene (Reduced Lateral Root Formation) is a pivotal genetic element in plant development, with functions that trace back to the earliest land plants. Initially identified in Arabidopsis thaliana for its role in lateral root formation, RLF encodes a cytochrome b5-like heme-binding protein. This protein is integral to early cell divisions during lateral root initiation and operates independently of the conventional auxin signalling pathways mediated by ARF7 and ARF19.[1.1, 2.1]
Recent research has expanded our understanding of RLF's evolutionary significance. A study published in New Phytologist revealed that RLF is also present in Marchantia polymorpha, a liverwort species that diverged early in plant evolution and lacks true roots. In Marchantia, RLF is essential for proper vegetative and reproductive organ development. Notably, functional interchangeability was demonstrated between the RLF genes of Arabidopsis and Marchantia, indicating a conserved role across diverse plant lineages.[3.1, 4.1]
The RLF protein's heme-binding capability suggests a link between metabolic processes and organ development. This discovery underscores the concept of exaptation in evolutionary biology, where existing genes acquire new functions over time. In this case, a gene originally involved in general organ development was co-opted for root formation in vascular plants.[5.1]
Understanding RLF's function provides insight into the molecular mechanisms underlying plant organogenesis and highlights the evolutionary strategies plants employ to develop new structures. Further exploration of RLF and its interactions may offer avenues for enhancing crop resilience and productivity through targeted manipulation of root and shoot architectures.[3.1]
A root development gene that’s older than root development
A gene that regulates the development of roots in vascular plants is also involved in the organ development of liverworts — land plants so old they don’t even have proper roots. The Kobe University discovery highlights the fundamental evolutionary dynamic of co-opting, evolving a mechanism first and adopting it for a different purpose later.
When scientists discover that a gene is necessary for the development of a trait, they are quick to ask since when this gene has been involved in this and how the evolution of the gene has contributed to the evolution of the trait.
My group previously discovered that a gene called RLF is necessary for lateral root development in the model plant Arabidopsis thaliana, but it was completely new that the group of genes RLF belongs to is involved in plant organ development. So we wanted to know whether the equivalent of this gene in other plants is also involved in similar processes.
Professor Fukaki Hidehiro, corresponding author
Department of Biology
Graduate School of Science
Kobe University, Kobe, Japan.
Fukaki turned to the simplest model land plant he could find, the umbrella liverwort Marchantia polymorpha. This plant doesn’t even have roots that can extract water or nutrients from the soil, but it has its own version of the RLF gene. Using this organism, the Kobe University plant biologist studied the function of its RLF gene and whether its function within the cell is comparable to the Arabidopsis version of the gene. In the journal New Phytologist, he and his team publish that liverworts lacking RLF have severe deformations in various organs, demonstrating that RLF is involved in organ development in these basic land plants as well. They could even show that the Arabidopsis gene could perform its function in the liverwort and the liverwort’s gene in Arabidopsis.
This shows that the two genes are functionally interchangeable as actors in organ development.
Professor Fukaki Hidehiro.
The RLF gene produces a protein that belongs to the vast group of heme-binding proteins and that means that it may bind a molecule called “heme,” which is involved in energy transfer within the cell.Given that heme-binding proteins were not known to be involved in organ development in plants, another significant point in our study is that we showed that the RLF protein actually has a heme, both in the liverwort and in Arabidopsis.
Professor Fukaki Hidehiro.
The Kobe University researcher expects that learning more about how the RLF protein interacts with others will clarify more about the evolution of plant organ development.
The fact that RLF plays an important role in organ development since at least the dawn of land plants is an example of how evolution often co-opts existing mechanisms for new functions, such as for root development, which evolved only after liverworts and mosses branched off the other land plants.
Professor Fukaki Hidehiro.
Publication:
The discovery of the RLF gene in both vascular plants and primitive liverworts provides compelling evidence for the evolutionary continuity of genetic mechanisms across the plant kingdom—something that poses significant problems for creationist claims. According to standard creationist narratives, complex features like roots are designed independently and uniquely for each "kind" of organism. Yet the presence of a shared, functionally conserved gene in both root-bearing and rootless plants contradicts the idea of independent, special creation.Summary
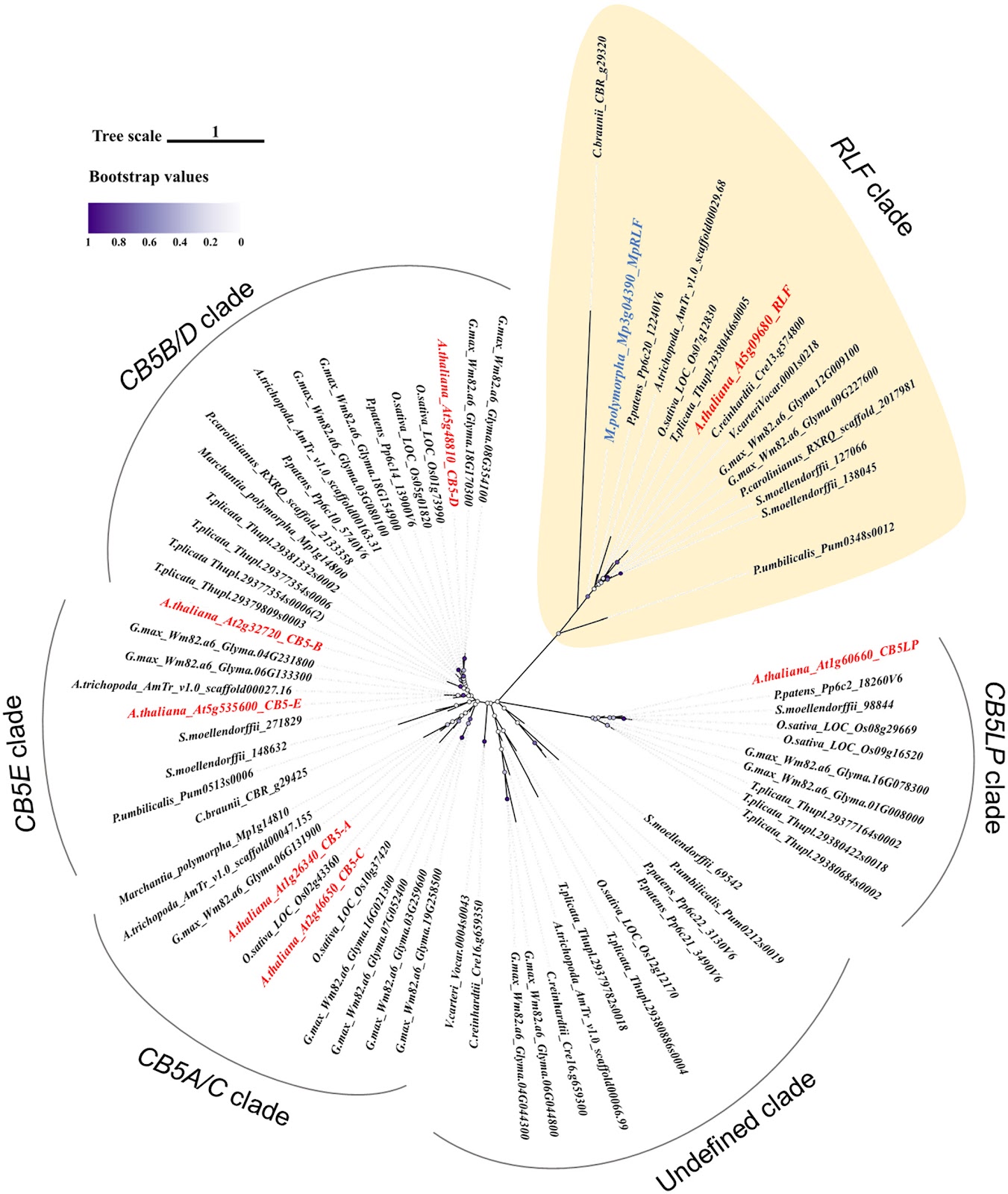
- In Arabidopsis thaliana, REDUCED LATERAL ROOT FORMATION (RLF), a cytochrome b5-like heme-binding domain (Cytb5-HBD) protein, is necessary for proper lateral root (LR) formation. Whereas the other Cytb5-HBD proteins in A. thaliana regulate different metabolic reactions, RLF is unique as it specifically regulates organ development. However, it remains unknown whether heme binding to RLF is necessary for its function and whether RLF orthologs in different plant species also regulate organ development.
- We demonstrate that RLF binds to heme in vitro and that two histidine residues, which are conserved among Cytb5-HBD, are crucial for both heme binding and its biological function in A. thaliana. In addition, we show that MpRLF, a RLF ortholog in the bryophyte Marchantia polymorpha, also binds to heme in vitro and that MpRLF rescues the LR formation phenotype of the A. thaliana rlf mutant.
- Mprlfge, the loss-of-function mutation in MpRLF, resulted in delayed thallus growth and inhibited both gemma cup and reproductive organ formation.
- Our findings indicate that MpRLF is essential for proper vegetative and reproductive development in M. polymorpha. This suggests that RLF-dependent redox reaction systems are conserved across diverse plant species and were independently co-opted for organ development in bryophyte and seed plant evolution.
Introduction
Heme is a porphyrin complex with a centrally coordinated iron atom. In plants, heme shares a metabolic pathway with Chl biosynthesis up to the production of protoporphyrin IX, after which ferrochelatase coordinates Fe2+ to protoporphyrin IX (Roper & Smith, 1997; Hederstedt, 2012). In the Protein Data Bank (https://www.rcsb.org), which contains 222 415 proteins, 4272 protein chains have been identified as heme-binding proteins (HBPs) from entries containing heme types b and c (Li et al., 2011). The binding of these HBPs to heme is maintained by the coordination of specific amino acids, such as histidine, within the apoprotein to the iron atoms of heme. In general, in various organisms, HBPs are essential for diverse biological processes such as steroid biosynthesis, aerobic respiration, and programmed cell death because of their role in electron transfer, substrate oxidation, and metal ion storage (Reedy & Gibney, 2004; Layer et al., 2010). In plants, HBPs, such as cytochrome c, which is involved in electron transfer during photosynthesis, and SOUL4, which is implicated in lipid metabolism within chloroplast fat droplets (plastoglobule), have demonstrated diverse physiological effects (Kerfeld & Krogmann, 1998; Shanmugabalaji et al., 2020). Recently, proteomic analyses in Arabidopsis thaliana have identified a variety of HBPs, such as basic/helix–loop–helix type nuclear transcription factors and intracellular signaling factors involved in GTPase activation (Shimizu et al., 2020.1). These studies have revealed that HBPs function as intracellular signals regulating diverse physiological functions. However, it remains unclear how they are involved in plant organ development.
One group of HBPs in plants is a family of proteins with a cytochrome b5-like heme-binding domain (Cytb5-HBD). In plants, the metabolic pathways mediated by most Cytb5-HBD proteins are diverse, including fatty acid desaturation, lignin biosynthesis, and nitrate reduction (Nagano et al., 2012.1; Gou et al., 2019). The genome of A. thaliana has 15 proteins with a Cytb5-HBD, including five members of the cytochrome b5 family (At1g26340 (CB5A), At2g32720 (CB5B), At2g46650 (CB5C), At5g48810 (CB5D), and At5g53560 (CB5E)) (Maggio et al., 2007), one cytochrome b5-like protein (At1g60660 (CB5LP)), four membrane-associated progesterone binding proteins (At2g24940 (MAPR2), At3g48890 (MAPR3), At4g14965 (MAPR4), and At5g52240 (MAPR5)) (Yang et al., 2005), two Δ-8 sphingolipid desaturases (At3g61580 (SLD1) and At2g46210 (SLD2)) (Sperling et al., 1998.1), two nitrate reductases (At1g77760 (NR1) and At1g37130 (NR2)) (Cheng et al., 1988; Wilkinson & Crawford, 1991) and At5g09680/REDUCED LATERAL ROOT FORMATION (RLF) (Ikeyama et al.,2010.1). In A. thaliana, cytochrome b5 isoforms, SLD1, SLD2, NIA1, NIA2, and proteins of the MAPR family generally do not play a significant role in organ development. CB5D is involved in lignin biosynthesis, and the cb5d mutant shows a reduction in S-lignin content. However, no abnormalities in organ development have been observed in cb5d mutants (Gou et al., 2019). SLD1 and SLD2 are involved in Δ-8 sphingolipid desaturation. Their loss-of-function mutants show significantly reduced root growth under sodium dodecyl sulfate (SDS) stress, but no growth defects are observed under normal conditions (Chen et al., 2012.2). Similarly, NIA1 and NIA2 are involved in the regulation of nitrate metabolism and nitric oxide (NO) production; however, the null-allelic mutant nia1-3 nia2-1 is lethal, and no studies have reported effects on organ development. Additionally, single mutants of either NIA1 or NIA2 do not exhibit significant effects on plant growth or development (Wilkinson & Crawford, 1993; Tang et al., 2022). Membrane-associated progesterone binding proteins, particularly MAPR5, contribute to lipid and steroid metabolism and regulate autophagy under starvation conditions, potentially influencing energy homeostasis. However, no severe developmental abnormalities have been reported in MAPR mutants or overexpression lines (Kimura et al., 2012.3; Ryu et al., 2017; Wu et al., 2021).
Whereas most of the Cytb5-HBD proteins in A. thaliana regulate a variety of metabolic reactions, At5g09680/RLF is a unique Cytb5-HBD protein that regulates organ development, including lateral root (LR) formation (Ikeyama et al., 2010.1). In general, LR formation is essential for root system architecture in most vascular plants and is regulated by auxin. In A. thaliana, LR formation is regulated by several auxin signaling modules including the SOLITARY-ROOT/IAA14, an auxin/indole-3-acetic acid (Aux/IAA) protein – AUXIN RESPONSE FACTOR (ARF) 7 and ARF19 module (Fukaki et al., 2005.1; Okushima et al., 2007.1). The rlf loss-of-function mutants of RLF show a marked reduction in the number of LRs while their ARF7/19-mediated auxin signaling is not affected, suggesting that RLF regulates LR formation independently of auxin response (Ikeyama et al., 2010.1). In addition, the rlf mutations cause aberrant organ development, including reduced primary root growth and decreased leaf size (Ikeyama et al., 2010.1). While the study of RLF in A. thaliana sheds light on the significance of Cytb5-HBD proteins for plant growth and development, a crucial gap remains in our understanding of how Cytb5-HBD proteins orchestrate growth and development of plant organs. First, it is not known whether RLF binds to heme and works as HBP in planta. Second, there is limited understanding of whether Cytb5-HBD proteins share a common role in regulating organ development in land plants. To address these issues, we extend our investigation of RLF to the model bryophyte species, Marchantia polymorpha (Ishizaki et al., 2016; Fernandez-Pozo et al., 2022.1) in addition to A. thaliana. M. polymorpha forms distinct organs specialized for vegetative or reproductive functions, such as the thallus, which serves as the main photosynthetic structure, gemma cups that produce gemmae, and reproductive organs like antheridiophores and archegoniophores. Its genetic tractability and evolutionary position as a nonvascular plant make it an ideal model for exploring these questions.
In this study, we demonstrate that the heme-binding ability of RLF is necessary for its biological function. In addition, we unveil the role of MpRLF, an ortholog of RLF in M. polymorpha, in orchestrating proper vegetative and reproductive development. Our findings highlight the crucial role of the MpRLF gene in plant organ development. Notably, the similarities between MpRLF and A. thaliana RLF suggest the existence of a conserved mechanism, governed by RLF that intricately regulates organ development across diverse plant lineages.
Iwata, K.P., Shimizu, T., Sakai, Y., Furuya, T., Fukumura, H., Kondo, Y., Masuda, T., Ishizaki, K. and Fukaki, H. (2025)
Evolutionary-conserved RLF, a cytochrome b5-like heme-binding protein, regulates organ development in Marchantia polymorpha. New Phytol. https://doi.org/10.1111/nph.70181
Copyright: © 2025 The authors.
Published by John Wiley & Sons, Inc. Open access.
Reprinted under a Creative Commons Attribution 4.0 International license (CC BY 4.0)
Instead, this finding is consistent with evolutionary theory, which predicts that new structures often evolve by repurposing existing genetic elements. In this case, a gene originally involved in general organ development in early land plants like Marchantia polymorpha has been co-opted and modified over evolutionary time to help regulate root development in vascular plants. This kind of genetic continuity—where genes acquire new functions while retaining traces of their ancestral roles—is a hallmark of descent with modification, not discrete acts of creation.
Furthermore, the fact that the Arabidopsis RLF gene can functionally substitute for the Marchantia version (and vice versa) despite hundreds of millions of years of evolutionary divergence indicates a deep genetic homology that is inexplicable under creationist models. Creationist arguments often dismiss such findings as examples of "common design," but this explanation fails to account for the imperfect, contingent, and historically layered nature of gene repurposing observed in living organisms.
Ultimately, the RLF gene's evolutionary history provides a textbook example of exaptation — a process by which evolution co-opts existing genes for new functions. Far from challenging evolutionary theory, this discovery adds yet another line of genetic evidence supporting the shared ancestry of all plant life and undermines the notion that each plant form was created in isolation with fully formed and uniquely designed features.








All titles available in paperback, hardcover, ebook for Kindle and audio format.
Prices correct at time of publication. for current prices.

No comments :
Post a Comment
Obscene, threatening or obnoxious messages, preaching, abuse and spam will be removed, as will anything by known Internet trolls and stalkers, by known sock-puppet accounts and anything not connected with the post,
A claim made without evidence can be dismissed without evidence. Remember: your opinion is not an established fact unless corroborated.