

Check any plan leaf and it will normally be one of two types - the monocotyledons with parallel leaf veins and the dicotyledons with a network of veins organised around a central vein with regular branches.
But this has not always been so. As a team of researchers at Vienna University, in collaboration with colleagues from the National Museum of Natural History in Stockholm and the Hebrew University in Jerusalem, have discovered, plants seem to have evolved the network pattern of leaf several times over the course of their evolution with most of them becoming extinct fairly quickly on an evolutionary timescale.
Their findings are published, open access, in the journal New Phytologist and explained in a University of Vienna news release:
Flowering plant-like leaf veins became extinct and re-evolved several times throughout Earth's historyTechnical detail and background to their research is given in the team's open access paper in New Phytologist:
According to a research team led by paleontologists from the University of Vienna, the reticulated leaf veins typical of flowering plants developed much earlier than expected, but died out several times. With the help of new methods, the fossil plant Furcula granulifer has now been identified as such a pioneer: the leaves of this seed fern species already showed the hierarchical, networked veining in the late Triassic (around 201 million years ago). The study was recently published in the journal New Phytologist .
Mario Coiro and Leyla Seyfullah from the Institute of Paleontology at the University of Vienna, in collaboration with colleagues from the National Museum of Natural History in Stockholm and the Hebrew University in Jerusalem, have discovered new clues to the origin of flowering plants - the most successful group of plants on earth:By looking through old collections, we were able to use new methods to identify a 201-million-year-old plant that developed similar characteristics to today's flowering plants - but without having the same evolutionary success: it became extinct.
Mario Coiro, co-corresponding author
Department of Palaeontology
University of Vienna, Vienna, Austria.Plant revolution through new leaf structureAlthough the 201-million-year-old fossil leaves of Furcula granulifer show the reticulated, hierarchical leaf venation typical of most modern plants, we found that Furcula actually belongs to the now extinct group of seed ferns. So it appears that this typical leaf venation, which enables efficient photosynthesis, has developed several times over the course of Earth's history.
Leyla J. Seyfullah, co-corresponding author
Head of the paleobotany and terrestrial paleoecology research group
Department of Palaeontology
University of Vienna, Vienna, Austria.
Flowering plants - more precisely, angiosperms - are the most important group of plants on Earth today, dominate most terrestrial ecosystems and are essential for human survival. Their appearance in the Cretaceous Period (approximately 145 - 66 million years before our time) revolutionized biodiversity on Earth and led to the spread of other groups such as mammals, insects and birds, thereby increasing the overall diversity on Earth.
This plant revolution was triggered in part by a novel leaf structure: the leaves of flowering plants have a network-like, hierarchical vein structure that can bind carbon dioxide much more efficiently. "Among the few other fossil plants with such leaf veins, the fossil leaf of Furcula granulifer from the Late Triassic of Greenland is particularly strikingly similar to today's structure - so much so that it was originally described as an angiosperm leaf, although according to current knowledge this group is only more so than 50 million years later," says Coiro. This caused debate in the scientific community, but the actual relationship of Furcula was not reanalyzed for almost 100 years.
See with new eyes
For the present study, the research team used historical material on the one hand, but also re-examined the samples using conventional and confocal laser scanning microscopy with regard to their microscopic structure and the structure of the layer that surrounds the leaves (cuticle). As a result, the Furcula could now be assigned to an extinct group of seed plants with fern-like leaves - "seed ferns" or pelatasperms.
The vein structure of the leaves has therefore developed independently several times in the course of evolution. It was also shown that the leaves of Furcula had a lower vein density - they were therefore not as efficient at binding carbon as today's flowering plants. Overall, it can therefore be viewed as a failed evolutionary experiment in the Late Triassic.
Failed evolution experiments
As another such failed evolutionary experiment, albeit around 50 to 100 million years earlier, the research team also identified another group of mysterious seed plants, the Gigantopteridales. They developed a similar leaf structure during the Permian period (approx. 300 million - 250 million years ago), but have also become extinct.Through these 'natural experiments' we have the opportunity to better understand the reasons for success and failure in the course of the evolution of flowering plants - the decisive factor is probably the combination of several characteristics rather than just a single key innovation.
Leyla J. Seyfullah
Creationists might like to speculate on why any omniscience, omnipotent intelligent designer would find several different ways of doing the same thing, most of which, apart from the last, failed and the species with them went extinct.SummaryIntroduction
- Leaf venation is a pivotal trait in the success of vascular plants. Whereas gymnosperms have single or sparsely branched parallel veins, angiosperms developed a hierarchical structure of veins that form a complex reticulum. Its physiological consequences are considered to have enabled angiosperms to dominate terrestrial ecosystems in the Late Cretaceous and Cenozoic.
- Although a hierarchical-reticulate venation also occurs in some groups of extinct seed plants, it is unclear whether these are stem relatives of angiosperms or have evolved these traits in parallel. Here, we re-examine the morphology of the enigmatic foliage taxon Furcula, a potential early Mesozoic angiosperm relative, and argue that its hierarchical vein network represents convergent evolution (in the Late Triassic) with flowering plants (which developed in the Early Cretaceous) based on details of vein architecture and the absence of angiosperm-like stomata and guard cells.
- We suggest that its nearest relatives are Peltaspermales similar to Scytophyllum and Vittaephyllum, the latter being a genus that originated during the Late Triassic (Carnian) and shares a hierarchical vein system with Furcula.
- We further suggest that the evolution of hierarchical venation systems in the early Permian, the Late Triassic, and the Early Cretaceous represent ‘natural experiments’ that might help resolve the selective pressures enabling this trait to evolve.
Leaves are crucial for the functioning of the Earth system, because they play a significant role in fundamental ecological processes, such as photosynthesis, transpiration, and nutrient cycling. Representing the primary site of photosynthesis, leaves are responsible for regulating the exchange of water vapour and other gases between plants and the atmosphere through their stomata, with direct consequences for the global hydrological cycle and for maintaining Earth's climate.
Venation is one of the most important regulators of leaf function, impacting not only hydraulics but also carbon investments, structure, rigidity, defences, and carbon assimilation (Sack & Scoffoni, 2013). Angiosperm leaves are characterised by a hierarchical venation system, with veins of different gauges branching from one another. Moreover, veins commonly merge together (anastomosis) and in many cases do not terminate at the margin, but instead within the leaf lamina. This system allows for extremely high vein density (de Boer et al., 2012), and impacts investment returns of carbon and nitrogen in the leaf (Zwieniecki & Boyce, 2014; Baresch et al., 2019). Thanks to these innovations, angiosperms achieve higher rates of transpiration, photosynthesis, and maximum carbon assimilation than those of other extant vascular plant clades (Brodribb et al., 2007; Boyce et al., 2009). The evolution of the angiosperms' unique venation system is among the key traits that enabled angiosperms to revolutionise terrestrial ecosystems through the Cretaceous (Benton et al., 2022).
Construction of cross-connections between veins requires additional investment in vascular tissue by the plant. However, reticulate venation provides clear benefits by increasing water and solute transport efficiency to and from the photosynthetic cells, increasing structural strength of the lamina, and improving the plant's capacity to divert resources around areas of physical or biological damage. As a consequence of these benefits, meshed venation systems appear to have evolved independently and at multiple times throughout the history of euphyllophytes (Alvin & Chaloner, 1970; Trivett & Pigg, 1996; Boyce & Knoll, 2002; Boyce, 2005). Other than in angiosperms, reticulate venation occurs in a broad range of plant groups including Ophioglossales, Marattiales, Polypodiales (Wagner, 1979), Medullosales (Zodrow et al., 2007.1), Glossopteridales (Pigg & McLoughlin, 1997), Peltaspermales (Krassilov, 1995), Bennettitales (Seward, 1903; Xu et al., 2023), Cycadales (Erdei & Manchester, 2015; Erdei et al., 2019.1; Coiro et al., 2023.1), Ginkgoales (Arnott, 1959), Petriellales (Bomfleur et al., 2014.1), Caytoniales (Halle, 1910), and Gnetales (Rydin et al., 2003; Yang et al., 2015.1; Coiro et al., 2022.1). Instead, venation combining a hierarchical structure with anastomosing venation is much less common, being present in some ferns (Wagner, 1979), Gigantopteridales (Glasspool et al., 2004), Gnetum L., and various seed plants of uncertain affinities (Cornet, 1993; Trivett & Pigg, 1996; Anderson & Anderson, 2003.1).
The Rhaetian (latest Triassic) was a Mesozoic greenhouse interval characterised by increasingly warm climates and high pCO2 (c. 1000 ppm background concentration) resulting in an absence of polar ice caps and highly diverse ecosystems with lush mid- and high-latitude vegetation (Kustatscher et al., 2018). During the Late Triassic, Greenland was part of a Northern American/Central European floral subprovince, which had many floristic elements, including Cycadales, Bennettitales, conifers, ginkgophytes, corystosperms and peltasperms, such as Lepidopteris Schimp., shared with floras across this broader region (Harris, 1926, 1937; McElwain et al., 2007.2; Kustatscher et al., 2018; Slodownik et al., 2021). The epoch terminated in the (mostly marine) end-Triassic mass extinction event, resulting in a floral turnover possibly caused by abrupt global warming and environmental degradation c. 201.3 million years ago (Ma) (Harris, 1937; Lundblad, 1959.1; McElwain et al., 1999, 2007.2; Steinthorsdottir et al., 2011, 2012.1, 2018.1).
Furcula, established by Harris (1932), is a rare component of the Greenland Late Triassic flora with unforked or once-forked leaves with each entire-margined lanceolate segment bearing a prominent midrib and fine reticulate subsidiary venation but lacking a continuous marginal vein. Harris (1932) also outlined various cuticular details and noted that the guard and subsidiary cells were apparently derived from the subdivision of a single mother cell. Based on its anastomosing venation and cuticular features, Furcula has long been considered a potential stem group of the angiosperm lineage (Scott et al., 1960; Maheshwari, 2007.3; Taylor et al., 2009.1). Indeed, Harris (1932) stated ‘If Furcula were a Tertiary fossil—particularly if only the undivided type of leaf were known—it would unquestionably be regarded as a Dicotyledon on the evidence of its venation alone’. However, Wolfe et al. (1975) noted that some of the characteristics of the vein anastomoses in Furcula (i.e. absence of terminal looping of the secondary veins) are fundamentally different to those in angiosperms, and Krassilov (2012.2) argued that the affinities of this genus are just as likely to lie with gigantopterids, peltasperms or gnetophytes.
Here, we reinvestigate Furcula based on new fossils and Harris' original material (macrofossils and cuticle preparations). We document the details of vein organisation, epidermal cell and stomatal arrangement, and opaque bodies within the mesophyll to evaluate characters shared with other seed plant groups. Our results not only shed new light on the botanical affinities of this peculiar forked leaf, but reveal extensive convergence towards high-density hierarchical vein systems during the Late Triassic. We discuss how the development and ultimate failure of this ‘natural experiment’ can inform our understanding of the success of the Cretaceous expansion of angiosperms and the importance of key traits in driving evolutionary radiations.
 Fig. 1.
Fig. 1.
Geographic and stratigraphic context of the Furcula granulifer fossils investigated in this study. Left: map of Greenland (azimuthal equidistant projection) with the locality of Scoresby Sound indicated with a black box. Right: Stratigraphic column of the section exposed in Jameson Land, with the main plant fossiliferous beds indicated (after Harris, 1937). The beds bearing fossils of Furcula granulifer are highlighted.
Fig. 2.
Furcula granulifer Specimen S167977. (a) Overview of the sample, showing three leaves (black arrows), with the leaves in a and b surrounded by ellipses; (b) detail of the upper leaf in a; (c) detail of the lower leaf in a, from which the cuticular samples were extracted. Bars: (a) 5 cm; (b, c) 1 cm.Fig. 3.
Macromorphology and venation of Furcula granulifer. (a) and (c) are details of slide 302 and slide 305, respectively, from Harris' preparations, with (b) and (d) being the isolated green channel from the image, shown to increase clarity. Images show the details of the venation and the anastomoses. Bars: (a) 1 cm; (c) 5 mm, (b) at same scale as a, (d) as same scale as c.Fig. 4.
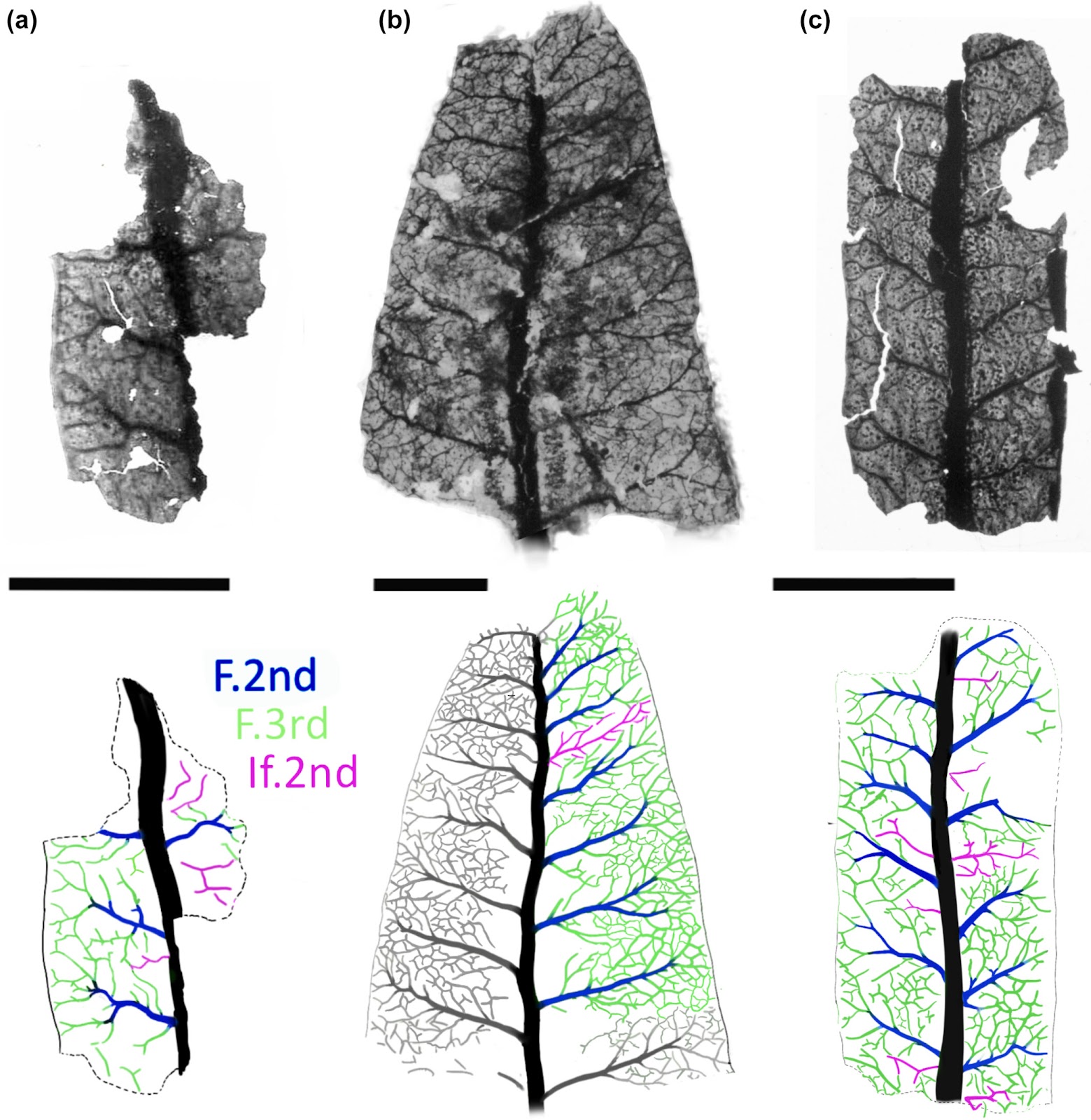
Venation plus diagrams interpreting the venation of Furcula granulifer. All images represent the green channel exported as grayscale with increased contrast. (a, c) Slide 309, (b) slide 302. The diagrams show the course of the fascicular secondaries (F.2nd), the fascicular tertiaries (F.3rd), and the interfascicular secondaries (If.2nd). Bars, 5 mm.Fig. 5.
Cuticular anatomy of Furcula granulifer specimen S167977. (a) Nonstomatiferous (adaxial) surface, observed directly on the specimen surface using a GFP filter. Mineral encrustations are present; (b) confocal maximum intensity projection of the nonstomatiferous surface, showing the presence of ‘quartet’ patterns; (c) sum of intensity projection of the stomatiferous (abaxial) surface, imaged directly on the coal; (d) inverted version of (c), more clearly showing the elongated pavement cells corresponding to veins (v) and the location of the stomata (st). (e) Isolated cuticle showing multiple stomata with papillae overarching the stomatal pit and the random arrangement of the guard cells. (f) stoma imaged on the coalified leaf surface, showing the radiating subsidiary cells ending in papillae. Bars: (a) 200 μm; (c) 100 μm; (b, d, e) 50 μm; (f) 10 μm.




But that might be easier for them than coping with the fact that the scientists show no signs of giving up on the Theory of Evolution and adopting creationism as a better explanation of the observable facts, like their cult leaders have fooled them into believing is about to happen, any day now, real soon and has been about to happen for about the last 20 years, during which a fresh generation of scientists have grown up and taken up research posts.
And then there is the fact that this all happened so long before there was an Earth for it to happen on, according the creationist superstition - something that doesn't ever seem to cause them the slightest doubt.
What Makes You So Special? From The Big Bang To You

Ten Reasons To Lose Faith: And Why You Are Better Off Without It


No comments :
Post a Comment
Obscene, threatening or obnoxious messages, preaching, abuse and spam will be removed, as will anything by known Internet trolls and stalkers, by known sock-puppet accounts and anything not connected with the post,
A claim made without evidence can be dismissed without evidence. Remember: your opinion is not an established fact unless corroborated.