

Artist's impression of the Ediacaran fauna
What creationists will tell you about the Cambrian 'Explosion', i.e., the rapid radiation of different species with different body plans, is that it happened in an instant. One minute there was nothing; the next, lots of different species with lots of different body plans were swimming about eating one another and avoiding being eaten. This is a parody of course and depends on deliberately or otherwise, misunderstanding what the term 'Cambrian Explosion' means.
What it doesn't mean is that lots of different body plans suddenly appeared without ancestor and with no common ancestry. Creationist claims of spontaneous, magical creation are nonsense, of course. Not only did the Cambrian biota have ancestors, but those ancestors themselves had ancestors in the preceding 'Ediacaran' era, named after the Australian rocks in which their fossils were first found. And those Ediacaran multicellular organisms had ancestors in the form of single-celled organisms that gave rise to these colonial organisms.
What caused the transition from the sessile Ediacaran biota to the Cambrian biota was the evolution of mobility. Mobility creates the opportunity for predation so, unlike sessile organisms that depend on a supply of simple nutrients that have to be metabolized and make into structural materials, being able to predate on other organisms means they can obtain organic molecules already made by something else.
What information is there on the transition from the Ediacaran to the Cambrian biota and the main environmental selectors that drove it? The transition from the Ediacaran to the Cambrian biota is one of the most significant events in the history of life on Earth, marked by the emergence of a wide variety of complex, multicellular organisms. This transition is often referred to as the "Cambrian Explosion," occurring approximately 541 million years ago. Here are key aspects of this transition and the main environmental selectors that drove it:And predation provides selection pressure to not only become more efficient predators than the competition, but it also provides selectors favouring defensive structures and strategies - like mobility, vision, burrowing, shells, spikes, jaws and means of grasping, and this is what we see developing in the Cambrian biota. What we also see emerging in the Cambrian is a bilateral body plan with a front and back end with the main sensory organs clustered at the front.
Characteristics of the Ediacaran Biota
- Time Period: The Ediacaran Period spans from about 635 to 541 million years ago.
- Organisms: Ediacaran biota included a range of simple, soft-bodied organisms. These organisms were often immobile or exhibited limited movement. They lacked hard parts, such as shells or exoskeletons.
- Examples: Notable Ediacaran organisms include Dickinsonia, Charnia, and Spriggina.
Characteristics of the Cambrian Biota
- Time Period: The Cambrian Period follows the Ediacaran, starting around 541 million years ago and lasting until about 485 million years ago.
- Organisms: The Cambrian Explosion saw the rapid diversification of life forms, including many that had hard parts like shells and exoskeletons. This period is characterized by the appearance of most major animal phyla.
- Examples: Notable Cambrian organisms include trilobites, brachiopods, and the enigmatic Hallucigenia.
Environmental Selectors Driving the Transition
Fossil Evidence and Research
- Increased Oxygen Levels
- Hypothesis: One of the primary drivers of the Cambrian Explosion is believed to be a significant increase in atmospheric and oceanic oxygen levels.
- Impact: Higher oxygen levels would have supported higher metabolic rates, larger body sizes, and the evolution of more complex body plans and behaviors.
- Changes in Ocean Chemistry
- Hypothesis: Changes in ocean chemistry, such as increased availability of calcium and other minerals, could have facilitated the development of hard parts like shells and skeletons.
- Impact: The ability to form hard parts provided advantages in protection from predators and structural support, leading to greater diversification.
- Predation Pressure
- Hypothesis: The emergence of new predatory species could have driven evolutionary arms races, leading to rapid diversification and the development of new defensive adaptations.
- Impact: Predation would have created strong selective pressures, favoring organisms with effective defenses and strategies for avoiding predators.
- Genetic and Developmental Innovations
- Hypothesis: Changes at the genetic and developmental levels, such as the evolution of new regulatory genes and pathways, could have enabled the development of more complex body plans.
- Impact: These genetic innovations would have provided the raw material for the evolution of diverse and complex life forms.
- Ecological Interactions and Niches
- Hypothesis: The establishment of complex ecosystems with diverse ecological interactions and niches could have promoted the rapid diversification of life.
- Impact: The availability of new ecological roles and interactions would have driven adaptive radiation, where organisms evolve to exploit different niches.
- Burgess Shale and Other Fossil Sites: The Burgess Shale in Canada, along with other fossil sites like the Chengjiang in China, provides critical evidence of the diverse Cambrian biota. These sites contain exceptionally well-preserved fossils that offer insights into early animal life.
- Molecular Studies: Advances in molecular biology and genetics have provided insights into the evolutionary relationships between Ediacaran and Cambrian organisms, helping to trace the development of complex life.
Conclusion
The transition from the Ediacaran to the Cambrian biota represents a profound shift in the history of life, driven by a combination of environmental changes, ecological interactions, and genetic innovations. The increase in oxygen levels, changes in ocean chemistry, the emergence of predation, and the development of complex ecosystems played crucial roles in this evolutionary milestone, leading to the rich diversity of life forms that characterize the Cambrian period.
Now three geobiologists from Edinburgh University's School of GeoScience, led by Dr. Fred Bowyer, have found evidence that major geological changes which caused sea-level changes, were also triggers for this transition because shallow water is more responsive to global atmospheric oxygen level changes. They have just published their findings, open access in the journal Science Advances and have explained it in an Edinburgh University press release:
Sea level changes shaped early life on Earth, fossils show
Shifts in the Earth’s continental plates that drove long-term changes in sea level set the stage for the evolution of the earliest animals on Earth, a study suggests.
A newly developed timeline of early animal fossils reveals a link between sea levels, changes in marine oxygen, and the appearance of the earliest ancestors of present-day animals.
The study reveals clues into the forces that drove the evolution of the earliest organisms, from which all major animal groups descended.
Ancient fossils
A team from the University of Edinburgh studied a compilation of rocks and fossils from the so-called Ediacaran-Cambrian interval – a slice of time 580–510 million years ago. This period witnessed an explosion of biodiversity according to fossil records, the causes of which have baffled scientists since Charles Darwin.
The early animals found from this era were all sea-dwellers, at a time when oxygen levels in the air and ocean were much lower than today.
While the very first lifeforms before this time were mostly single-cell, and simple multi-celled organisms, creatures in the Ediacaran Period started to become more complex, with multiple cells organised into body plans that allowed them to feed, reproduce, and move across the ocean floor.
This era also marked the emergence of so-called bilaterian animals – which display symmetrical body plans, in common with most present-day species including humans.
Biodiversity timeline
By compiling data from different sources – including radioactive dating and geochemical information about the layers of rock in which fossils were found – the team mapped all major fossil finds and various environmental datasets onto a single timeline.
The new chronology allowed the team to study trends in biodiversity for the period in question with more detail than before.
They combined these insights with further chemical clues from the geological record – confirming a link between major changes in global sea levels, intervals when shallow marine environments gained more oxygen, and the appearance and diversification of early animal groups.
This dynamic set the stage for several significant bursts in biological diversity, known as the Avalon, White Sea, and Cambrian assemblages, each marking the arrival of new animal groups and the decline of others.
By reconstructing environmental conditions in deepest time, the study unlocks new insights into the ancient forces and pressures that shaped the earliest life on our planet.
The team also identified gaps in the fossil record, suggesting that current knowledge about early animals is biased by the clusters of sites worldwide where fossils have been found and studied.
Constructing a timescale of early animal evolution using the rock record is a daunting task, only made possible through international and interdisciplinary research. But an integrated global approach is crucial. It exposes biases in our records, while also revealing patterns in fossil appearances, sea level cycles, and environmental oxygen.
Dr Fred Bowyer, lead author
School of Geosciences Edinburgh University, Edinburgh, UK
Knowing what drives biodiversity is a fundamental piece of knowledge in the puzzle of life. I feel very privileged to have built upon decades of interdisciplinary global research, and contributed to a better understanding of the role that sea level plays in early animal evolution.
Mariana Yilales Agelvis, co-author
School of Geosciences Edinburgh University, Edinburgh, UK
The paper was published in Science Advances and funded by the Natural Environment Research Council (NERC).
Abstract
The drivers of Ediacaran-Cambrian metazoan radiations remain unclear, as does the fidelity of the record. We use a global age framework [580–510 million years (Ma) ago] to estimate changes in marine sedimentary rock volume and area, reconstructed biodiversity (mean genus richness), and sampling intensity, integrated with carbonate carbon isotopes (δ13Ccarb) and global redox data [carbonate Uranium isotopes (δ238Ucarb)]. Sampling intensity correlates with overall mean reconstructed biodiversity >535 Ma ago, while second-order (~10–80 Ma) global transgressive-regressive cycles controlled the distribution of different marine sedimentary rocks. The temporal distribution of the Avalon assemblage is partly controlled by the temporally and spatially limited record of deep-marine siliciclastic rocks. Each successive rise of metazoan morphogroups that define the Avalon, White Sea, and Cambrian assemblages appears to coincide with global shallow marine oxygenation events at δ13Ccarb maxima, which precede major sea level transgressions. While the record of biodiversity is biased, early metazoan radiations and oxygenation events are linked to major sea level cycles.
INTRODUCTION
The Ediacaran-Cambrian appearance and rise of animals (metazoans) from approximately 575 to 515 Ma are seminal events in evolution. However, the underlying processes and drivers of this diversification, such as the relative influence of extrinsic, environmental factors versus intrinsic species interactions, remain unresolved. To interrogate this event, we first need to establish the global record of biodiversity itself.
The modern distribution of biodiversity is spatially highly heterogeneous, as biodiversity is distinctly regionalized, with high diversity in the tropics and a large fraction hosted in a limited number of environments or ecological communities [e.g., (1)]. Larger geographic areas also have higher diversities than smaller areas, known as the species-area effect (2). It is expected, then, that the spatial variation in biodiversity at any given moment in deep time has been shown to be greater than the temporal variation for most of the Phanerozoic fossil record of marine metazoans (3). Interactions between species and their environments can influence speciation and extinction rates within any given region, but over geological timescales, global diversity can also be a function of evolutionary innovation, global tectonic and climatic drivers (which may regulate global provinciality), as well as changes in atmospheric composition, biogeochemical cycling, and oceanographic circulation, although these control biodiversity via local- or regional-scale processes (4).
However, the fossil record of biotic diversity changes cannot be read literally. The record is notoriously incomplete and subject to many biases of sampling and preservation. The degree to which the fossil record has been explored, researched, and documented also varies, biasing the recording of data. In addition, Konservat-Lagerstätten provide exceptional windows into local biotas at episodic times and places during the history of life, and the early to middle Cambrian is famously replete with these horizons that have yielded some of the best-studied biotas of the fossil record [e.g., (5, 6)]. Conditions for this exceptional preservation are created by the local dynamics of physical sedimentary deposition, as well as favorable redox profiles and early diagenetic environments, which are, in part, controlled by global seawater chemistry and oxygenation.
Changes in habitable marine shelf area, where most early metazoans lived, are controlled by relative sea level rise (transgression) and fall (regression), which, in turn, also drives the long-term geological record of marine sedimentary rocks [e.g., (7)], and there is a well-documented covariation between Phanerozoic metazoan macroevolutionary patterns and sedimentary rock availability [e.g., (8)]. This suggests that expansion of marine shelf environments coincided with the diversification of animals via the species-area effect (2). The late Ediacaran to Cambrian system is globally characterized by extensive continental flooding over the Great Unconformity, accompanied by a notable increase in preserved rock area/volume dominated by shallow marine settings (9). A strong correlation between marine sedimentary area and volume flux has been confirmed for the North American Ediacaran-Cambrian rock and metazoan fossil record, although, here, the marked rise of Cambrian reconstructed biodiversity cannot be explained by the increase in Cambrian sedimentary volume alone (10). These analyses suggest that there may be some secular bias in the rock record through this interval (11). However, while this correlation may indicate preservational biases, it also suggests that there are shared underlying driving processes that control both an increase in reconstructed biodiversity and preserved sedimentary rock quantity, such as expressed in the interrelationships between sea level, the carbon cycle, nutrient delivery, and changing oceanic redox state [e.g., see (12–15)].
Although the distribution of fossil metazoan taxa is highly variable temporally and spatially (4, 16–18), the Ediacaran-Cambrian transition is commonly viewed as comprising a succession of four metazoan radiations (Fig. 1). The Ediacara biota (~575 to ≤538 Ma) may include possible nonmetazoans [e.g., (19)] and stem-group and early crown-group eumetazoans [e.g., (20)]. The bilaterian body plan was probably present by ~560 Ma, followed by a marked increase in bilaterian fossils in the early Cambrian (the Cambrian “Explosion”) by which time most phyla had appeared (21). Phanerozoic evolutionary faunas are broadly defined by high-level taxonomic groups and display a broadly synchronous logarithmic increase in reconstructed biodiversity followed by extinctions, with the increase in diversity associated with successive faunas coinciding with a decline of the previously dominant fauna (8). The Ediacara biota has been similarly subdivided into three temporally overlapping assemblages (or paleocommunities) (Fig. 1B). These, however, are thought to have occupied different sedimentary settings: The soft-bodied Avalon assemblage (rangeomorphs, arboreomorphs, and nonfrondose morphogroups) initially occupied deeper marine siliciclastic settings, but representatives are also found in mid-depth carbonate and siliciclastic settings <560 Ma; the soft-bodied White Sea assemblage (erniettomorphs, dickinsoniomorphs, triradialomorphs, penta/octoradialomorphs, bilateralomorphs, kimberellomorphs, and problematica) occupied shallow to mid-depth siliciclastic marine settings; and the soft-bodied and skeletal (calcified) Nama paleocommunity (dominated by cloudinomorphs) is found in dominantly shallow marine siliciclastic and carbonate settings (16, 17, 22–24).Apparent extinctions, as well as possible ecological shifts, are suggested to have shaped early metazoan evolutionary dynamics (17, 25). The Avalon assemblage appears very broadly coincident with positive δ13Ccarb values that predate the onset of the Shuram δ13Ccarb excursion [Fig. 1; e.g., (26)]. Both the Avalon and White Sea paleocommunities largely disappeared by ~550.5 Ma (25, 27, 28), but the causes are still unclear, largely because of uncertainties in global chemostratigraphic correlation between approximately 560 and 550 Ma. The first calcified metazoans appear ~550.5 Ma in shallow marine carbonate rocks globally (18, 29) and increase in diversity substantially in the Cambrian, mainly as small shelly fossils (SSFs) found within shallow marine carbonates. The Ediacaran-Cambrian boundary, defined by the first appearance of the ichnospecies Treptichnus pedum and the lowest level of the associated T. pedum ichnozone, has been suggested to coincide with, or immediately postdate, the nadir of the “Basal Cambrian negative carbon isotope (δ13C) excursion” (BACE), but both the precise position of this first appearance relative to the BACE nadir and the age of onset of the BACE (~538 to 535 Ma) remain uncertain (18, 30). This is due, in part, to a global paucity of continuous Ediacaran-Cambrian successions. The disappearance of the Ediacara biota and subsequent radiation of the Cambrian biota have also been hypothesized to result from the decline of unique taphonomic preservational modes, an absence of outcrops of appropriate sedimentary settings, or a mass extinction [e.g., (31, 32)]. This biotic transition broadly coincides with the BACE, which marks a global perturbation of the marine carbon cycle [e.g., see (30, 33–35)], but the relationship of this to any potential extinction mechanism, such as a rise in shallow marine anoxia, is not established. The four Ediacaran to early Cambrian paleocommunities therefore differ in their preferred depositional settings, and so our ability to confidently track evolutionary change over time may be hampered by the uneven or potentially biased temporal and spatial distribution of their respective rock records. Fig. 1. Integrated chronology and biostratigraphy of globally distributed Ediacaran to earliest Cambrian successions.
Fig. 1. Integrated chronology and biostratigraphy of globally distributed Ediacaran to earliest Cambrian successions.
(A) Radiometric ages (horizontal lines; table S1) calibrate the 10-point moving average of an updated global composite δ13Ccarb database [data S1 and S2 updated after (18, 28, 30)]. Gray-shaded envelope shows uncertainty in the δ13Ccarb framework associated with data scatter. (B) Compilation of estimated ranges of major Ediacaran to earliest Cambrian (580 to 528 Ma) inferred metazoan morphogroups and major trace fossil groups within the Avalon, White Sea, Nama, and Cambrian assemblages, showing dominant sedimentary setting of host successions. Age ranges, uncertainties, and full references for biostratigraphic database are provided in data S1. Modified from (18, 24). Question marks denote taxa with the most uncertain maximum/minimum age constraints. See fig. S2 for an expanded version of this figure. SPIE, Spitskop excursion; OME, Omkyk excursion; WSI, White Sea Interval; BAINCE, Baiguoyuan negative carbon isotope excursion.
Calibrated molecular clock studies show that the origin of the Metazoa, the last common eumetazoan ancestor, the protostome-deuterostome ancestor, and all bilaterian clades predate the first appearance of macroscopic representatives in the fossil record (36). This is not only due to issues of preservation, sampling, and acquisition of taxonomically identifiable features but also suggests that developmental and morphological novelties, as well as the unique characters that define the origin of major metazoan groups, may be decoupled from the drivers that allowed them to gain large body size, proliferate, and hence rise to ecological prominence in the fossil record, i.e., features that promote the expression as radiations in the fossil record. Intervals when metazoans dominate and radiate therefore often record the operation of ecological or extrinsic drivers (36).
Evolutionary innovations can also create dynamic feedbacks of species interactions, as they may act as ecosystem engineers and alter the flow and structural pathways of energy through the biosphere (4). The diversification of bilaterian animal phyla in the Cambrian must have increased metazoan abundance and fueled interspecific competition, thereby further facilitating biodiversification and biomineralization and promoting the rise of more energy intensive ecologies, such as burrowing and predation, leading to escalatory arms races [e.g., see (37)]. Some have further suggested that the early radiation of bilaterians may have caused the extinction of the Ediacaran biota by slow biotic replacement [see review in (32)].
Changes in global shallow marine oxygen levels during the Ediacaran-Cambrian interval have been inferred on the basis of interpretations of paleoredox proxy data, including uranium isotopes (δ238U). Seawater δ238U compilations from both carbonate and siliciclastic rocks suggest that multiple oscillations in global marine redox conditions occurred at 1- to 10-Ma timescales [e.g., (38–40)]. Available geochemical data therefore support recent modeling efforts (41), which show that any increase in environmental oxygen levels during the Ediacaran-Cambrian interval did not proceed unidirectionally but rather via a series of oxygenation pulses that likely correspond with atmospheric and “oceanic oxygenation events” [OOEs; e.g., (42)]. These OOEs would likely have been accompanied by corresponding regional and/or global changes in nutrient and productivity regimes and may have been driven, in part, by changing weathering dynamics, including changes in the composition of weathering material [e.g., (42, 43)].
The assumption that δ238U data can be used as a proxy to track changes in global open ocean paleoredox conditions has been challenged by studies, which show that sea level changes [e.g., (44)], as well as local productivity and basin restriction [e.g., (45)], also control δ238U variability in the sedimentary record. In particular, a decline in δ238U has been noted to coincide with local relative sea level fall and changes in primary carbonate mineralogy and diagenesis (44), and the offset in δ238U between seawater and mudstone records has been shown to be sensitive to basin hydrography, organic carbon export dynamics, and changes in sedimentation rate and lithology (45). Careful screening of geochemical data, however, combined with an integrated multiproxy approach that considers sea level change, regional basin dynamics and associated degrees of restriction, and global chronostratigraphic constraints and associated uncertainties, has the potential to reveal redox-related trends. Integrating δ238Ucarb and carbonate-associated sulfate (δ34SCAS) isotopic data reveals that supposed OOEs in the Ediacaran-Cambrian interval occur synchronously with changes to the global marine carbon cycle, as indicated by co-occurring trends in δ13Ccarb data from the same samples, in addition to trends in coeval data revealed by regional and global chemostratigraphic correlation (39, 46, 47). This implies that broad trends in data compilations of multi-isotope systems (e.g., δ13Ccarb─δ238Ucarb─δ34SCAS) track a common process, which may reflect the extent of anoxic seafloor that coincides with high SO4 reduction rates and pyrite and organic carbon burial rates. In this model, the onset of decreasing δ13Ccarb and δ34SCAS has been interpreted to reflect the onset of shallow marine oxygenation (46). Decreasing δ13Ccarb and δ34SCAS (falling limbs) reflects progressive oxygenation of the deeper water column, resulting in decreasing organic carbon and pyrite burial, which is consistent with coeval increases in baseline δ238Ucarb data. This decreased reductant burial would gradually slow/stall atmospheric oxygenation (46). Increasing δ13Ccarb and δ34SCAS (rising limbs) and δ238Ucarb falling limbs then captures marine deoxygenation conducive to the progressive burial of organic carbon and pyrite, driving a long-term build-up of atmospheric oxygen (39, 46). Falling and rising “limbs” of the C─S─U isotope systems across this time interval may thereby reflect the balance between marine and atmospheric oxygen dynamics that control, and are controlled by, the relative areal extent of marine conditions conducive to the burial of organic carbon and pyrite (48). The area of seafloor available to host organic carbon and pyrite burial in productive regions of the open ocean, in turn, should be controlled by changes in relative sea level and productivity, both of which are also influenced, to some degree, by global plate reorganization and associated mid-ocean ridge length, eustatic sea level change, and prevailing regional and global climate and associated weathering regimes (43). Regional differences in absolute magnitude of isotopic data (e.g., δ13Ccarb) during globally recognized excursions have also been suggested to reflect regional depositional and early diagenetic regimes [e.g., (49)], and alternative drivers for Neoproterozoic δ13Ccarb negative excursions have also been proposed to reflect changes in the areal extent of shallow marine carbonate settings conducive to photosynthetic primary production and local isotopic reservoir effects [e.g., (50)]. Each of these mechanisms is strongly influenced by eustatic and relative sea level change.
A causal relationship between shallow marine oxygenation and the Cambrian radiation has long been proposed [e.g., (51)], but it remains unclear as to how rising oxygen availability, as well as possible associated changes in nutrient regimes, may have promoted early animal radiations. Increasing atmospheric oxygen concentration may have deepened the redoxcline, so extending habitable water depths, promoted animal-sediment mixing [e.g., (52)], and allowed for the evolution of more metabolically costly ecologies such as carnivory, in turn driving a predator-prey “arms race” and hence diversification (53). Spatially and temporally dynamic shallow marine anoxia itself might have formed physical barriers to dispersal, thereby promoting reproductive isolation and speciation (54, 55). This is inferred from the early Cambrian record of the Siberian Platform, which shows that phases of increased diversity of reef biota coincide with a deepening of the habitable zone on the shallow marine shelf which, in turn, is interpreted to correspond with a possible deepening of the redoxcline (56). These phases also coincide with increased speciation during local sea level lowstands, and it is suggested that lowstand intervals permitted extensive oxygenation of shallow waters over the entire craton, thereby providing oxic corridors for dispersal and the creation of new founder communities (55).
A paucity of globally distributed radiometric constraints due to limited datable ash beds, and numerous biostratigraphic correlation uncertainties, have hindered evaluation of metazoan distribution, sedimentary rock volume, and environmental change throughout the Ediacaran-Cambrian interval. Here, we use a global age framework for the interval from 580 to 510 Ma interval, that temporally calibrates available data from 24 regional composite successions (Figs. 2 and 3 and fig. S1). The fully integrated age framework construction considers all available high-precision radiometric ages, δ13Ccarb chemostratigraphy, δ238Ucarb data, and the biostratigraphic distribution of metazoan genera [updated after (18); tables S1 and S2, data S1 to S4, and figs. S1 to S6]. We use this to reconstruct mean genus richness and include all known Ediacaran Lagerstätten but exclude all early Cambrian examples up to 510 Ma from our compiled diversity data [for full list, see (6)], of which the most biodiverse are the Kuanchuanpu, Chengjiang, Sirius Passet, Sinsk, and Emu Bay Shale biotas (Fig. 3). Hence, our Cambrian reconstructed biodiversity data consist of the skeletal metazoan record only. This framework assumes an onset age for the BACE at ~535 Ma and an age of ~533 Ma for the first appearance of T. pedum after (30) (see Material and Methods, Figs. 1 and 3A, fig. S2, tables S1 and S2, and data S1 to S4). To reduce uncertainty, only successions where there is some confidence in temporal global correlation and spatial extent throughout the target interval are included (see the Supplementary Materials for expanded discussion and fig. S1). Hence, this record represents only a variable fraction of total global rock quantity in any one region. While absolute values of, e.g., calculated volume flux and area may therefore have limited utility, long-term temporal trends are likely to be globally meaningful and will capture underlying mechanistic processes (see data S3). These data therefore allow investigation of this interval in three ways. First, we estimate temporal changes in the global marine sedimentary rock volume and area together with the distribution of different sedimentary settings to explore how the biotic record is controlled by the availability of host rocks (see Material and Methods for definition of different sedimentary settings). Preserved sedimentary volume flux (area × thickness) is mainly controlled by accommodation (governed by relative sea level fluctuations) and sediment supply, while area, which might theoretically be a more reliable metric of changing available habitable sea floor through time, is also governed by sea level transgression/regression. Both metrics might relate to habitability and so both are considered. We test the correlation between the reconstructed biodiversity, rock volume/area, and sampling intensity, using Spearman’s rank correlation (which tests for monotonic linear and nonlinear relationships). Second, we investigate the potential mechanism of long-term sea level change as a driver for observed carbon cycle dynamics and environmental oxygenation using the calibrated records of δ13Ccarb and δ238Ucarb. Third, we test the hypothesis that global redox changes controlled successive radiations and extinctions through the Ediacaran-Cambrian transition. Fig. 2. Terrane/outcrop area (maximum/minimum/approximate) calculated for each region investigated.
Fig. 2. Terrane/outcrop area (maximum/minimum/approximate) calculated for each region investigated.
See table S2 and data S3 for full data.
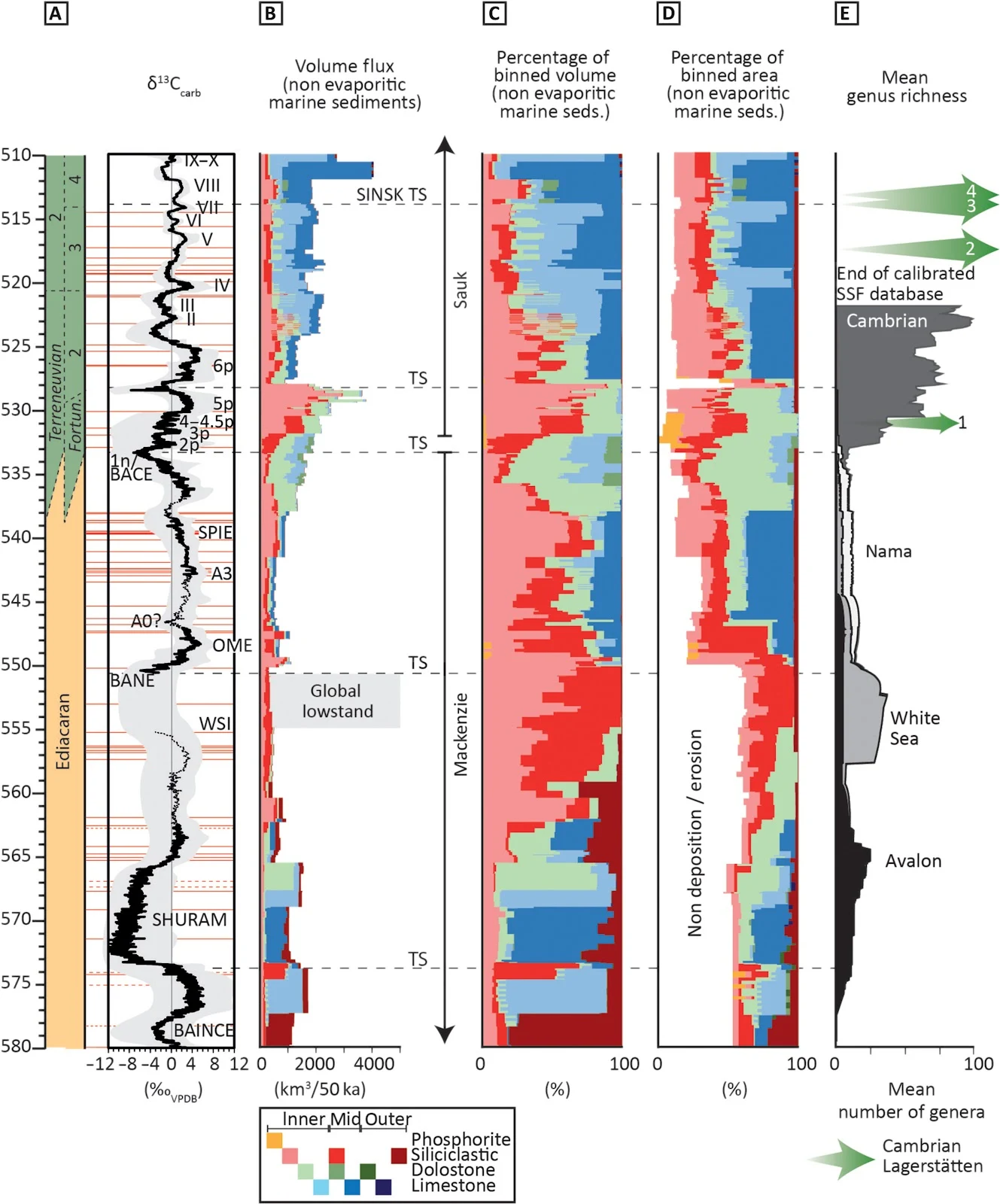 Fig. 3. Compilation of trends in marine sedimentary rocks and hiatuses from 580 to 510 Ma.
Fig. 3. Compilation of trends in marine sedimentary rocks and hiatuses from 580 to 510 Ma.
(A) Radiometric ages (horizontal red lines; table S1) calibrate the 10-point moving average of an updated global composite δ13Ccarb curve, with associated uncertainty [data S1 and S2 updated after (18, 28, 30)]. VPDB, Vienna Pee Dee belemnite. (B) Estimated volume flux of globally distributed marine sedimentary rocks (table S2 and data S3). (C) Percentage of total volume of nonevaporitic marine sedimentary rocks (in each 50-ka bin). TS, transgressive surface. (D) Percentage of total area of nonevaporitic marine sedimentary rocks, including proportion of record represented by nondeposition or erosion (in each 50-ka bin). (E) Reconstructed biodiversity (mean genus richness) organized into four successive assemblages based on constituent morphogroups, after (24). Example Cambrian Lagerstätten: 1, Kuanchuanpu; 2, Chengjiang; 3, Sirius Passet; 4, Sinsk and Emu Bay Shale.
Fred T. Bowyer et al.
Sea level controls on Ediacaran-Cambrian animal radiations.
Sci. Adv. 10, eado6462 (2024). DOI:10.1126/sciadv.ado6462
Copyright: © 2024 The authors.
Published by American Association for the Advancement of Science. Open access.
Reprinted under a Creative Commons Attribution 4.0 International license (CC BY 4.0)



Just as the Theory of Evolution predicts, environmental change maps onto changes in biodiversity through an evolutionary process. The geological evidence confirms both in the fossil record and the evidence of oxygenation levels.
To anyone who understands the subject, this will come as no surprise at all since the geological record is a record of what actually happened. The fact that it refutes creationist is because creationism never happened so it left no record. For some reason, creationists seem unable to make that link between what the record shows, what actually happened and what they believe happened. Creationism is nothing if not a delusional detachment from reality.
What Makes You So Special? From The Big Bang To You

How did you come to be here, now? This books takes you from the Big Bang to the evolution of modern humans and the history of human cultures, showing that science is an adventure of discovery and a source of limitless wonder, giving us richer and more rewarding appreciation of the phenomenal privilege of merely being alive and able to begin to understand it all.
Available in Hardcover, Paperback or ebook for Kindle
Ten Reasons To Lose Faith: And Why You Are Better Off Without It

This book explains why faith is a fallacy and serves no useful purpose other than providing an excuse for pretending to know things that are unknown. It also explains how losing faith liberates former sufferers from fear, delusion and the control of others, freeing them to see the world in a different light, to recognise the injustices that religions cause and to accept people for who they are, not which group they happened to be born in. A society based on atheist, Humanist principles would be a less divided, more inclusive, more peaceful society and one more appreciative of the one opportunity that life gives us to enjoy and wonder at the world we live in.
Available in Hardcover, Paperback or ebook for Kindle

No comments :
Post a Comment
Obscene, threatening or obnoxious messages, preaching, abuse and spam will be removed, as will anything by known Internet trolls and stalkers, by known sock-puppet accounts and anything not connected with the post,
A claim made without evidence can be dismissed without evidence. Remember: your opinion is not an established fact unless corroborated.