Figure 1.
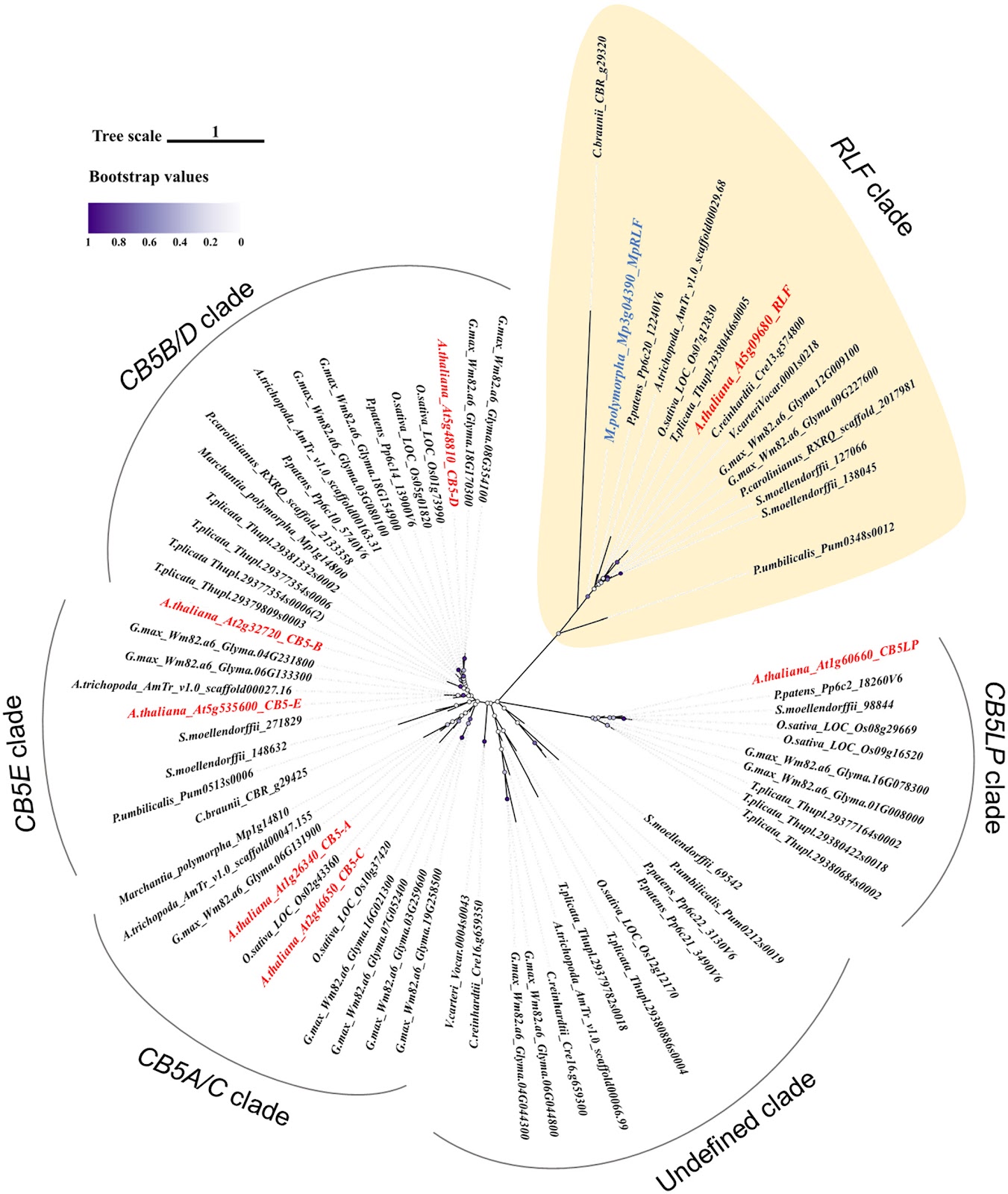
Foraging of urediniospores by bees on plants infected with myrtle rust. a. Honey bee forager collecting Austropuccinia psidii urediniospores from leaves of broadleaf paperbark [Melaleuca quinquenervia (Myrtales, Myrtaceae)], Bungawalbin, New South Wales, Australia; b. A. psidii urediniospores in the opening of a Geraldton wax [Chamelaucium uncinatum (Myrtales, Myrtaceae)] flower bud in Brisbane, Queensland, Australia.
Foraging of urediniospores by bees on plants infected with myrtle rust. a. Honey bee forager collecting Austropuccinia psidii urediniospores from leaves of broadleaf paperbark [Melaleuca quinquenervia (Myrtales, Myrtaceae)], Bungawalbin, New South Wales, Australia; b. A. psidii urediniospores in the opening of a Geraldton wax [Chamelaucium uncinatum (Myrtales, Myrtaceae)] flower bud in Brisbane, Queensland, Australia.
Photographs by Geoff Pegg.
A paper published in March 2026 in NeoBiota is entirely unsurprising to an evolutionary biologist, but deeply awkward for anyone trying to present nature as the intelligent design of an omnibenevolent creator. It reports what appears to be a mutually beneficial relationship between the introduced Western honey bee, Apis mellifera, and the invasive fungal plant pathogen myrtle rust, Austropuccinia psidii. It is a neat example of how evolution has no foresight, no moral purpose and no long-term plan. Symbiotic alliances can arise naturally between different species when there is an immediate benefit to both, even when the longer-term consequences for one or both partners — and for the wider ecosystem — may be destructive.
Myrtle rust, Austropuccinia psidii, is an invasive rust fungus in Australia. It infects more than 500 species in the Myrtaceae family, which includes many of Australia’s culturally, ecologically and economically important native plants, including eucalypts, paperbarks and related species. In Australia, the pathogen is regarded as a serious threat to native ecosystems, with around 17% of endemic vegetation considered at risk. The other partner in this apparent mutualism is also an introduced species: the Western honey bee, Apis mellifera.
The basis of this relationship is an exquisite example of the sort of functional complexity creationists routinely try to claim as evidence for intelligent design. That, of course, raises the obvious question: why would an omnibenevolent designer design a fungal pathogen capable of damaging so much of Australia’s native vegetation, and then provide it with a convenient pollinator-assisted dispersal system?
The mechanism is ingenious, but only in the blind, short-term sense in which natural selection can be ingenious. The bright yellow urediniospores of myrtle rust are collected by honey bees much as pollen is collected. The bees pack the spores into their pollen baskets and carry them back to the hive. For the fungus, this provides a potential route for dispersal beyond simple wind movement. For the bees, the spores are not just inert particles accidentally mistaken for pollen; they have real nutritional value. The researchers found that myrtle rust spores contained more than 22% protein and all ten amino acids regarded as essential for honey bee nutrition, making them comparable with high-quality pollen.
Laboratory feeding trials also showed that honey bee larvae could develop normally on a diet based on myrtle rust spores, with survival, development time and body weight similar to larvae reared on a high-quality pollen diet. Even more concerning from a biosecurity point of view, the spores remained viable inside honey bee colonies for at least nine days, meaning that hives could potentially act as reservoirs and transport systems for the pathogen.
So, the fungus gains a mobile vector, while the bees gain an alternative protein source. But what benefits both in the short term could be damaging in the longer term. As myrtle rust damages Myrtaceae-rich habitats, it can reduce the availability of flowers and pollen. That, in turn, could encourage honey bees to rely more heavily on fungal spores or other alternative foods, while the fungus benefits from the continued movement of bees and managed hives. The result could be a damaging ecological feedback loop, with plant-pollinator networks and forest regeneration placed under increasing pressure.
This is precisely the kind of outcome that any omniscient, intelligent designer should have foreseen and avoided. It is also precisely the kind of outcome that a mindless evolutionary process can produce without difficulty. Natural selection rewards immediate reproductive or nutritional advantage; it does not plan for ecological stability, protect biodiversity, or ensure that mutually beneficial relationships remain harmless in the long term.