This post is a bit of a change from my usual fare, where I’m busy pulling apart the pseudo-science of creationism with its reliance on magic and one ancient book of myths from a small Bronze Age tribe. Today, instead, I want to share something more uplifting: a glimpse of the rich and colourful life thriving in Britain’s coastal waters.
The footage comes from the Isles of Scilly, a protected area lying just off the tip of Cornwall in the far southwest of Britain. Thanks to the Gulf Stream, these islands enjoy a climate that feels almost Mediterranean — a rare treat for the UK!
A team from the University of Exeter’s Centre for Ecology and Conservation set up remote cameras around the islands to see what they could capture. This was a proof-of-concept project designed to test new ways of monitoring coastal ecosystems. The results didn’t disappoint. Their findings have just been published open-access in the Ecological Society of America’s journal Ecological Applications.
Artist’s reconstruction of Badjcinus timfaulkneri scavenging on the carcass of Silvabestius michaelbirti, a wombat-like marsupial also from Hiatus Site.
88 years ago today, the last Tasmanian tiger died in Beaumaris Zoo, Hobart, Tasmania.
It was the last in a line of carnivorous marsupial going back at least 23-25 million years ago, in the extended period of Earth's pre-'Creation Week' history when creationists believe there was nothing out of which a magic man made of nothing made everything in 6 days, just 10,000 years ago. According to creationist mythology, the Tasmanian tiger would have had no ancestors, being magically made from dirt just like every other living species.
This makes it hard for creation to explain the evidence of all those fossils known to be far older than 10,000 years and showing evidence of the evolution of extant species from ancient ancestors as species diversified from a single common ancestor which lived hundreds of millions of years ago, but then facts were never a problem for counter-factual creationism.
What information do you have on the extinct Tasmanian tiger?
The Tasmanian tiger, also known as the thylacine (Thylacinus cynocephalus), was a carnivorous marsupial native to Tasmania, mainland Australia, and New Guinea. The thylacine looked somewhat like a medium-to-large dog with a slender body, a head resembling that of a wolf or dog, and distinctive dark stripes across its back and tail, giving it the nickname "Tasmanian tiger." However, it was more closely related to kangaroos and other marsupials.
Key Information about the Tasmanian Tiger:
Physical Description:
The thylacine was about the size of a large dog, with a length of up to 1.8 to 2 meters (including the tail) and a height at the shoulder of about 60 cm.
Its fur was short and coarse, typically tan or yellow-brown, with 13 to 20 dark stripes across its back and tail.
It had a long, stiff tail, similar to a kangaroo's, and could open its jaws incredibly wide, up to 120 degrees.
Habitat and Behavior:
The thylacine was a nocturnal and solitary animal, hunting at night and resting during the day in a den or hollow log.
It primarily fed on small to medium-sized animals, such as kangaroos, birds, and small mammals. Its diet may have also included carrion.
Extinction:
The thylacine was driven to extinction on the Australian mainland thousands of years ago, likely due to competition with humans and the dingo, a wild dog introduced to Australia.
In Tasmania, the species persisted until the 20th century. The last known wild thylacine was killed in 1930, and the last known individual in captivity died in 1936 at the Hobart Zoo in Tasmania.
Its extinction was primarily due to human activities, including hunting (encouraged by a bounty system), habitat destruction, and the introduction of diseases.
Conservation Efforts and "Rediscovery" Claims:
Despite being declared extinct, there have been numerous unconfirmed sightings and efforts to find surviving thylacines, but none have been scientifically verified.
Some scientific projects have explored the possibility of "de-extinction" using preserved DNA from museum specimens, but these efforts remain largely theoretical.
Cultural Significance:
The thylacine holds a unique place in Tasmanian culture and Australian folklore, often seen as a symbol of the tragedy of extinction and the need for wildlife conservation.
It appears on Tasmania's coat of arms and has been the subject of various books, documentaries, and studies.
Would you like to know more about any specific aspect of the Tasmanian tiger?
How the oldest ancestors of the Tasmanian tiger were discovered by palaeontologists from the University of New South Wales, Australia, is the subject of a Taylor & Francis news release:
Earliest known ancestors of the extinct Tasmanian Tiger discovered – and some could even eat the bones and teeth of their preyA marsupial which had an “extremely thick” jawbone, enabling it to consume even the bones and teeth of its prey is among a discovery of three, new ancient species of the modern Thylacines – otherwise known as the Tasmanian tiger, which went extinct 88 years ago.
These new species each roamed Australia around 23-to-25 million years ago, during the late Oligocene, making them the “undoubted oldest members of this family ever discovered”.
Today – Australia’s National Threatened Species Day, which marks the death of the last Tasmanian Tiger in Hobart’s Beaumaris Zoo on the 7th of September 1936 – scientists from the University of New South Wales (UNSW) Vertebrate Palaeontology Lab publish their findings in the Journal of Vertebrate Paleontology.
The once suggested idea that Australia was dominated by reptilian carnivores during these 25 million-year-long intervals is steadily being dismantled as the fossil record of marsupial carnivores, such as these new thylacinids, increases with each new discovery. The diversity of mammalian carnivores at Riversleigh during this period rivals that seen in any other ecosystem, including the great mammalian carnivore radiation that developed in South America.
Timothy Churchill, lead author
University of New South Wales
Sydney, New South Wales, Australia.
The three new species were each found in the fossil-rich deposits in Riversleigh World Heritage Area.
The largest of these new species, Badjcinus timfaulkneri, weighed somewhere between 7-11 kilograms, about the same size as a large Tasmanian Devil. Alike the Tasmanian Devil, timfaulkneri possessed an extremely thick jawbone enabling it to consume the bones and teeth of its prey. This species is related to the much smaller, previously discovered B. turnbulli (2.7 kg) – which until now was the only other undoubted thylacinid known from the late Oligocene.
The dentary and isolated first molar of B. timfaulkneri were recovered from Hiatus Site which is even older than Riversleigh’s White Hunter Site where B. turnbulli was previously found, making B. timfaulkneri the oldest undoubted thylacine discovered so far.
Badjcinus timfaulkneri is named after Tim Faulkner, the director and co-owner of the Australian Reptile Park and managing director of Aussie Ark. Tim has dedicated his life to the conservation of Australia’s wildlife including the largest still-living marsupial carnivore, the Tasmanian Devil.
The second new species is Nimbacinus peterbridgei. This was about the size of a Maltese Terrier (~3.7 kg). This species is represented by a near-complete dentary from White Hunter Site. Nimbacinus peterbridgei was a predator that probably focused on small mammals and other diverse prey species that lived with it in the ancient forests. Species of Nimbacinus appear to be more closely related to the Tasmanian Tiger than other thylacinids of similar age. This means Nimbacinus peterbridgei is probably the oldest direct ancestor of the Tasmanian Tiger yet known.
Nimbacinus peterbridgei was named after Australian geologist, speleologist and bibliophile Peter Bridge. He has devoted his life to helping uncover Australia’s ancient past, particularly in the caves of Western Australia.
The last species, Ngamalacinus nigelmarveni was a ~5.1kg thylacinid – approximately the size of a Red fox. It was also from White Hunter Site at Riversleigh. The blades on the lower molars of species of Ngamalacinus are elongated with deep V-shaped carnassial (‘meat-cutting’) notches, suggesting they were highly carnivorous – more so than any of the other thylacinids of similar size.
Ngamalacinus nigelmarveni is named after Nigel Marven, a renowned British television documentary presenter famous for paleontology-inspired series like Prehistoric Park and Sea Monsters.
The presence of three distinct lineages of specialised thylacinids during the late Oligocene highlights how quickly they diversified after first appearing in the fossil record. These thylacinids exhibits very different dental adaptations, suggesting there were several unique carnivorous niches available during this period. All but one of these lineages, the one that led to the modern Thylacine, became extinct around 8 million years ago.
That lineage of these creatures that survived for more than 25 million years ended with the death of Benjamin, the last Tasmanian Tiger in Hobart’s Beaumaris Zoo on the 7th of September 1936.
Professor Michael Archer, co-author
University of New South Wales
Sydney, New South Wales, Australia.
ABSTRACT
New thylacinid species of Badjcinus, Nimbacinus, and Ngamalacinus are described from upper Oligocene deposits of the Riversleigh World Heritage Area, northwestern Queensland. Badjcinus timfaulkneri, Nimbacinus peterbridgei, and Ngamalacinus nigelmarveni are among the oldest thylacinids yet known and indicate an earlier diversification of the family than previously understood. Maximum parsimony analysis supports a sister group relationship between Ng. nigelmarveni and Ng. timmulvaneyi, but the relationships of the two other new taxa are unresolved. Bayesian dated total evidence analysis using morphological and molecular data supports the generic assignment of B. timfaulkneri and Ng. nigelmarveni but not that of Ni. peterbridgei. Both phylogenies herein support a taxonomic reassignment of Thylacinus macknessi to the genus Wabulacinus, a conclusion also supported by the results of previous studies. Body mass estimates based on molar size regressions indicate body sizes ranging from 3.7 kg to 11.4 kg for the new thylacinid species. Badjcinus timfaulkneri exhibits an extremely deep jaw compared with other thylacinids, with mandibular bending strength analysis suggesting that it was a highly durophagous carnivore much like the modern dasyurid Sarcophilus harrisii. This analysis also suggests Ni. peterbridgei had a dentary more similar in shape to that of plesiomorphic thylacinid faunivores such as Ni. dicksoni and T. cynocephalus suggesting that it had a relatively more generalist faunivorous diet. The molars of Ng. nigelmarveni suggest they were better suited for longitudinal slicing than the molars of B. timfaulkneri and Ni. peterbridgei, indicating a more hypercarnivorous diet compared with that of those species.
INTRODUCTION
There are 12 extinct species in the dasyuromorphian family Thylacinidae, 10 of which are from the Oligo-Miocene (26–5.3 Ma) (Rovinsky et al., 2019). Unlike during the Plio-Pleistocene (5.3–0.12 Ma), which is dominated by large hypercarnivorous species of Thylacinus (15–55 kg), the Oligo-Miocene radiation exhibits considerably higher generic diversity, with seven monospecific clades known from the Riversleigh World Heritage Area in northwestern Queensland, all of which were between 3–10 kg in body mass. This restriction in size suggests thylacinids occupied most of the small- to medium-sized faunivorous niches at Riversleigh (1–10 kg). Contemporary peramelemorphians (∼50 g–1.5 kg) (Gurovich et al., 2014; Travouillon et al., 2013; Travouillon et al., 2010, 2014.1) and thylacoleonids (∼10–50 kg) (Gillespie, 2023; Gillespie et al., 2016, 2019.1a, 2019.2b) occupied relatively smaller and larger carnivore niches.
The oldest undoubted thylacinid, Badjcinus turnbulli Muirhead & Wroe, 1998, is known only from White Hunter Site in the Riversleigh World Heritage Area. This and other late Oligocene sites at Riversleigh have not yet been radiometrically dated. White Hunter Site is interpreted to be late Oligocene in age because of the presence of the ilariid Kuterintja ngama, otherwise only known from the Ngama Local Fauna from the Etadunna Formation, which is magnetostratigraphically dated to 24.8–25 Ma (Myers & Archer, 1997; Woodburne et al., 1994). Riversleigh’s Faunal Zone A (FZA) deposits contain taxa that support a late Oligocene age (Arena et al., 2016.1; Travouillon et al., 2006). The relatively plesiomorphic dentition of B. turnbulli has led to difficulty in taxonomic assignment, with phylogenetic analyses placing it either as a basally branching member of Thylacinidae (Kealy & Beck, 2017; Muirhead & Wroe, 1998; Murray & Megirian, 2006.1a; Wroe & Musser, 2001), as a stem dasyurid (Wroe et al., 2000), or as a sister group to Dasyuromorphia as a whole (Kealy & Beck, 2017).
Only two other thylacinid fossils are known from upper Oligocene deposits. Originally assigned to Nimbacinus dicksoni but later reassigned to Thylacinidae incertae sedis, an isolated m2 (QM F16809) from D-Site at Riversleigh is the only other thylacinid known from Riversleigh’s upper Oligocene deposits (Muirhead & Archer, 1989; Murray & Megirian, 2000.1; Wroe & Musser, 2001). An isolated, broken M2 (NTM P2815–10) of a thylacinid is also known from the Pwerte Marnte Marnte Local Fauna (LF) in the Northern Territory, a deposit assumed to be upper Oligocene on the basis of biocorrelation (Murray & Megirian, 2006.2b; Woodburne et al., 1994). Although this upper molar, which is similar in size to the putative thylacinid Mutpuracinus archibaldi, was suggested by Murray and Megirian (2006.2b) to be the oldest thylacinid in the fossil record, there are doubts about its identification as a thylacinid (see Discussion).
Nimbacinus dicksoni Muirhead & Archer, 1989, is the best preserved and researched Miocene thylacinid (Attard et al., 2014.2; Murray & Megirian, 2000.1; Wroe & Musser, 2001). Multiple specimens including a near complete skull and skeleton are known from Middle Miocene deposits at Riversleigh and from the Bullock Creek LF in the Northern Territory. The dentition of Ni. dicksoni is relatively more plesiomorphic than that of other thylacinids (except possibly Muribacinus gadiyuli) in retaining slightly reduced metaconids on m2–4 and unreduced stylar cusps on M1–3. Previous morphofunctional analyses of the skull of Ni. dicksoni suggest it was a voracious predator capable of hunting prey larger than itself, with biting capabilities most similar to extant species of Dasyurus, rather than to the larger Thylacinus cynocephalus (Attard et al., 2014.2).
In addition to Nimbacinus dicksoni, four monotypic thylacinid genera are known from upper and lower dentitions recovered from Early and Middle Miocene Riversleigh deposits. These include the medium-sized (∼5–7 kg) Wabulacinus ridei and Ngamalacinus timmulvaneyi Muirhead, 1997.1, as well as the diminutive (∼1–2 kg) Muribacinus gadiyuli Wroe, 1996, and the large (∼18 kg) Maximucinus muirheadae Wroe, 2001.1a (Myers, 2001.2). The dentition of Ng. timmulvaneyi, W. ridei, and Ma. muirheadae have been considered to be relatively plesiomorphic in comparison with species of Thylacinus, but more derived than Ni. dicksoni (Muirhead, 1997.1). The diminutive Mur. gadiyuli is dentally the most plesiomorphic thylacinid known (Wroe, 1996).
Thylacinus macknessi Muirhead, 1992, from Riversleigh’s Early Miocene Neville’s Garden Site, is tentatively regarded as the earliest known member of the genus Thylacinus (Muirhead & Gillespie, 1995). This taxon possesses unique dental adaptations associated with a shift within the Thylacinus lineage towards hypercarnivory, including near complete loss of metaconids on m2–4, loss of entoconids, reduction of the lingual portion of the talonid margin in m2–4 and a lingual shift of the hypoconid so that the cristid obliqua forms a continuous longitudinal blade with the paracristid.
The powerful thylacine, T. potens Woodburne, 1967, from the Alcoota Local Fauna in the Northern Territory, has been interpreted on the basis of biocorrelation to be Late Miocene between 8.5 and 5.5 Ma (Megirian et al., 1996.1, 2010.1). It is the largest and most hypercarnivorous thylacinid known. It has been estimated to be between 30–56 kg in adult body mass (Myers, 2001.2; Wroe, 2001.1a), with some estimates exceeding 120 kg (Yates, 2014.3). Two additional species of Thylacinus with dental adaptations for increased carnivory, T. yorkellus and T. megiriani, are known from Upper Miocene to Lower Pliocene deposits (Murray, 1997.2; Yates, 2015).
Tyarrpecinus rothi Murray & Megirian, 2000.1, is a small thylacinid from the Late Miocene Alcoota Local Fauna of the Northern Territory. It is known from a broken maxilla that retains P2 and an isolated M2. The phylogenetic relationships and paleobiology of this taxon cannot be confidently determined until more complete craniodental material is found.
The quoll-sized Mutpuracinus archibaldi Murray & Megirian, 2000.1, known from a near complete skull with an incomplete upper and lower dentition from the Middle Miocene Bullock Creek LF in the Northern Territory (Murray & Megirian, 2006.1a), was initially considered to be an early thylacinid because of craniodental and basicranial similarities to plesiomorphic thylacinids such as Ni. dicksoni. However, more recent phylogenetic analyses suggest it should be regarded as Dasyuromorphia incertae sedis because of its lack of craniodental synapomorphies uniting it with undoubted thylacinids (Churchill et al., 2023.1; Kealy & Beck, 2017; Rovinsky et al., 2019).
Two further medium-sized (1–10 kg) incertae sedis dasyuromorphians are known from Miocene deposits; Whollydooleya tomnpatrichorum Archer, Christmas et al., 2016.2, from Miocene deposits in New Riversleigh (an area approximately 10 km southwest of the Riversleigh World Heritage Area) and Apoktesis cuspis Campbell, 1976, from upper Oligocene deposits at Lake Ngapakaldi in the Tirari Desert of South Australia. Either may be related to thylacinids or dasyurids (Archer, Christmas et al., 2016.2; Campbell, 1976). However, the lack of adequate fossil material to critically assess the relationships of these two enigmatic taxa precludes a more precise assignment beyond Dasyuromorphia incertae sedis. Whollydooleya tomnpatrichorum is only known from a single lower molar while the location of the skull and dentaries attributed to A. cuspis is currently unknown.
In this study, we describe three new species of thylacinid from Riversleigh’s oldest deposits: the upper Oligocene Hiatus and White Hunter Sites. The fossil specimens herein are all dentaries retaining near complete or partial lower dentitions.
Of course, it would be stupid to assume the Bronze Age pastoralists who wrote the Bible knew anything about Australia, or even a southern hemisphere. Since they believe Earth was a small flat place with a dome over it, they wouldn't even have considered hemispheres, northern of southern. Their view of the world was so narrow and restricted they thought all they had to explain was the small area within a day or two's walk of the Canaanite Hills, so nothing outside that small area was included; not an animals, mountain, continent or people; nothing.
So, the idea that they would have believed the Tasmanian tiger was magically created without ancestors 10,000 years ago or less is utterly preposterous and could only be believed by someone at least as blind to the evidence as were the authors of Genesis.
News that a team of evolutionary biologists are reassessing what we thought we knew about the dodo, Raphus cucullatus, is a good excuse to look at what the dodo teaches us about evolution.
The dodo was a species of pigeon which had spread to the Indian Ocean island of Mauritius, where in common with many island birds had lost the ability to fly. In the context of an island with no mammalian predators, any advantage of flight is balanced against the greater disadvantage of occasionally being blown out to sea. Natural selection, with no foresight and no plan, results in a flightless bird, and, as events transpire, extinction.
The arrival of Europeans with their rats, cats and dogs and need for supplies for a long sea voyage, was an environmental change that the dodo could not adapt to because evolution has no reverse gear and rarely reinvents what was lost earlier. The ground-nesting dodo like its close relative, on a neighbouring island, the Rodriguez Island Solitaire, Pezophaps solitaria, were doomed. They were extinct by the late 17th century.
Evolution is an unintelligent process. This would not be so, if species like the dodo had been intelligently designed by an omniscient designer, who could have foreseen and prepared for the coming of Europeans with their rats, cats and dogs. It is not an intelligent act to design a species headed for inevitable extinction because it can't adapt to what can be anticipated.
What information do you have on the 'Oxford dodo' and it's role in literature and biology?
The "Oxford dodo" is a significant artifact in both biological and literary history. Here's an overview of its importance:
Historical Background
The Dodo: The dodo (Raphus cucullatus) was a flightless bird native to Mauritius, an island in the Indian Ocean. It became extinct in the late 17th century due to overhunting by humans and the introduction of non-native animals.
Oxford Dodo: The Oxford dodo refers to the most complete known specimen of the dodo, which is housed at the Oxford University Museum of Natural History. This specimen includes the only remaining soft tissue (a dried head and a foot) from any dodo in the world.
Role in Biology
Scientific Study: The Oxford dodo has played a crucial role in scientific studies on the species. Since it includes soft tissue, researchers have been able to extract DNA, providing insights into the dodo's genetic makeup and its relationship to other bird species.
Extinction Studies: The dodo is often cited as one of the earliest recorded examples of human-caused extinction. The Oxford specimen has contributed to studies on extinction, illustrating the impact of human activity on isolated ecosystems.
Role in Literature
Lewis Carroll's "Alice's Adventures in Wonderland": One of the most famous literary references to the dodo is in Lewis Carroll's 1865 novel *Alice's Adventures in Wonderland*. The dodo in the story is believed to represent Carroll himself, who had a stammer and sometimes introduced himself as "Do-do-Dodgson" (Carroll's real name was Charles Dodgson). The character of the dodo in the story is depicted as a wise figure who proposes the "Caucus-race," where everyone wins and all must have prizes—a satire on political processes.
Symbol of Extinction: In literature and popular culture, the dodo has come to symbolize obsolescence and extinction, often representing something that is outdated or doomed to disappear. The Oxford dodo, as a physical reminder of a lost species, has reinforced this symbolism.
Cultural and Educational Impact
Exhibitions: The Oxford dodo has been featured in various exhibitions, not just as a scientific specimen but also as a cultural icon. It serves as a powerful educational tool about biodiversity loss and conservation.
Inspiration for Research and Art: The dodo has inspired a wide range of research, from studies in paleontology and conservation biology to its portrayal in art, literature, and even in discussions about colonialism and its impact on indigenous species.
Recent Developments
DNA Analysis: Advances in DNA technology have allowed scientists to conduct more detailed studies on the Oxford dodo's remains, leading to a better understanding of its biology and the causes of its extinction. These studies are crucial for understanding the dynamics of extinction and the evolution of island species.
Conservation Lessons: The story of the dodo, as illustrated by the Oxford specimen, is frequently used in discussions about conservation and the need to protect endangered species today. It serves as a cautionary tale about the consequences of human activity on the environment.
In summary, the Oxford dodo is more than just a biological specimen; it is a symbol of extinction and a cultural icon that has influenced literature, conservation science, and our understanding of human impact on the natural world.
Is there any evidence for the story that it was rescued from a bonfire of 'rubbish' at either the Ashmolean or the Oxford University Natural History Museum?
The story that the Oxford dodo was rescued from a bonfire of "rubbish" at the Ashmolean Museum, or the Oxford University Museum of Natural History is a widely circulated anecdote, but its accuracy is somewhat debated. Here's what is known:
Origins of the Story
Historical Context: The story typically claims that the dodo specimen, or parts of it, were nearly discarded during a clean-up or reorganization at one of the Oxford museums, likely the Ashmolean Museum, where the dodo was initially housed. The account suggests that someone recognized the value of the remains just in time to save them from being burned as rubbish.
Attribution: This anecdote is often linked to John Tradescant the Younger, an early naturalist and collector, whose collection formed the core of the Ashmolean Museum's early holdings. It is also sometimes associated with Elias Ashmole, the museum's founder.
Supporting Evidence
Lack of Concrete Records: There is no definitive archival evidence or documentation to substantiate the bonfire rescue story. The tale seems to have been passed down through oral history and later written accounts, but without solid documentary support.
Museum Records: Some records suggest that by the late 18th century, parts of the dodo specimen were discarded because they had decayed. The surviving head and foot were kept, while the rest of the body, which had likely decomposed significantly, was disposed of. This might have contributed to the myth that the specimen was almost entirely lost.
Plausibility and Scholarly Opinions
Scholarly Views: Historians and museum curators generally agree that while the story is plausible, it is likely an exaggeration or a simplification of events. It is plausible that during a period of reorganization or decay of the specimen, parts of the dodo were discarded, and someone recognized the importance of preserving the head and foot.
Interpretation: The bonfire story has become part of the dodo's lore, serving as a dramatic reminder of how close we came to losing this invaluable specimen. Whether or not it is literally true, the story underscores the precarious nature of historical artifacts and the importance of preservation.
Cultural Impact
Moral of the Story: The bonfire rescue tale is often used to highlight the fragility of historical artifacts and the importance of vigilance in preservation. It also serves to enhance the mystique of the dodo, a species already surrounded by a sense of loss and extinction.
In summary, while the story of the Oxford dodo being rescued from a bonfire at either the Ashmolean or the Oxford University Museum of Natural History is an intriguing one, it remains an anecdote with limited concrete evidence. The story has likely been embellished over time, though it reflects the real dangers historical specimens face when not properly cared for.
The extinction of the dodo on Mauritius had a long-lasting effect on the island's ecosystem of which the dodo was an essential part, but its contribution to the history of Oxford and it's literary heritage that is perhaps as interesting, coming as it does from the fact that the 'New' Ashmolean Museum became the depository of the only known remains of a dodo, which, when the collection in the Ashmolean was shared with the Oxford University Natural History Museum, and Pitt Rivers museum, was housed there, where it fascinated the Oxford Academic, Charles Ludwig Dodgson, aka Lewis Carroll (the Latinized form of Charles Ludwig).
Dodgson had a stammer and would introduce himself as Charles Do-Do-Dodgson, earning him the nickname Dodo Dodgson. When he wrote Alice in Wonderland, he based the Dodo on the 'Oxford dodo' and himself.
Part of the Oxford story concerning the Oxford dodo is probably apocryphal but serves to illustrate the importance of conservation, the fragility of specimens and recognising the importance of specimens which may not seem important at the time. The story is that it was saved from destruction at the last minute when it was destined for a bonfire of 'rubbish' that was being thrown out. The story varies as to who rescued it and whether it was the Ashmolean or the Natural History Museum that was about to burn it. What is probably true, and probably contributed to the story, is the fact that there was initially much more of the dodo but much of it decayed and was destroyed, leaving only the beak, feet and skin of the head and face.
The new study by researchers from the University of Southampton and Oxford University Museum is the subject of an open access paper in the Zoological Journal of the Linnean Society, and a University of Southampton News release:
Rethinking the Dodo Researchers are setting out to challenge our misconceptions about the Dodo, one of the most well-known but poorly understood species of bird.
In a paper published today in the Zoological Journal of the Linnean Society researchers from the University of Southampton, Natural History Museum (NHM) and Oxford University Museum of Natural History have undertaken the most comprehensive review of the taxonomy of the Dodo and its closest relative, the Rodriguez Island Solitaire.
They’ve painstakingly gone through 400 years’ worth of scientific literature and visited collections around the UK to ensure this iconic species, embodying humanity’s destructive potential, is correctly classified.
The Dodo was the first living thing that was recorded as being present and then disappeared. Before this, it hadn’t been thought possible for human beings to influence God’s creation in such a way. This was a time before the scientific principles and systems we rely on to label and classify a species were in place. Both the Dodo and the Solitaire were gone before we had a chance to understand what we were looking at.
Dr Neil J. Gostling, co-corresponding author
School of Biological Sciences
Faculty of Environmental and Life Sciences
University of Southampton, Southampton, UK.
Correcting the record
Much of what was written about the Dodo and the Solitaire was based on accounts from Dutch sailors, representations by artists, and incomplete remains.
The lack of a definitive reference point (type specimen) or convention to label species (zoological nomenclature) led to a series of misidentifications in the centuries following their extinction. New species such as the Nazarene Dodo, the White Dodo, and the White Solitaire were named, but the paper confirms that none of these creatures existed. Still, these erroneous ‘pebbles’ sent ripples through the waters of zoological literature.
By the 18th and early 19th centuries, the Dodo and the Solitaire were considered to be mythological beasts. It was the hard work of Victorian-era scientists who finally proved that the Dodo and the Solitaire were not mythological but were giant ground doves. Unfortunately, no one could agree how many species there had been. Throughout most of the 19th and 20th centuries, researchers thought there were three different species, although some people thought there had been four or even five different species.
Dr. Mark T. Young, co-corresponding author
School of Biological Sciences
Faculty of Environmental and Life Sciences
University of Southampton, Southampton, UK.
To unpick this confusion, researchers went through all the literature on the Dodo and Rodriguez Solitaire encompassing hundreds of accounts dating back to 1598 and visited specimens around the UK, including the world’s only surviving soft tissue from the Dodo, in the Oxford Museum.
More has been written about the Dodo than any other bird, yet virtually nothing is known about it in life. Based on centuries of nomenclatural confusion, and some 400 years after its extinction, the Dodo and Solitaire, continue to prompt heated debate. We’ve gone from where the first statements were made, seen how these have developed, and identified various rabbit holes to correct the record, as best we can.
Dr Julian Hume, co-author
Bird Group
Natural History Museum
Tring Hertfordshire, UK.
Through this work, researchers were able to confirm that both birds were members of the columbid (pigeon and dove) family.
Understanding its wider relationships with other pigeons is of taxonomic importance, but from the perspective of conservation, the loss of the dodo and the solitaire a few decades later means a unique branch of the pigeon family tree was lost. There are no other birds alive today like these two species of giant ground dove.
Dr Neil J. Gostling.
Challenging our misconceptions
The Dodo was an integral part of the ecosystem of Mauritius.
Artwork by Julian Pender Hume.
The researchers believe the popular idea of the Dodo as a fat, slow animal, predestined for extinction is flawed.
Even four centuries later, we have so much to learn about these remarkable birds. Was the Dodo really the dumb, slow animal we’ve been brought up to believe it was? The few written accounts of live Dodos say it was a fast-moving animal that loved the forest.
Dr. Mark T. Young.
Evidence from bone specimens suggests that the Dodo’s tendon which closed its toes was exceptionally powerful, analogous to climbing and running birds alive today. The dodo was almost certainly a very active, very fast animal. These creatures were perfectly adapted to their environment, but the islands they lived on lacked mammalian predators. So, when humans arrived, bringing rats, cats, and pigs, the Dodo and the Solitaire never stood a chance.
Dodos held an integral place in their ecosystems. If we understand them, we might be able to support ecosystem recovery in Mauritius, perhaps starting to undo the damage that began with the arrival of humans nearly half a millennium ago.
Dr Neil J. Gostling.
Learning ‘valuable lessons’
The study marks the beginning of a wider project to understand the biology of these iconic animals.
The mystery of the Dodo bird is about to be cracked wide open. We have assembled a fantastic team of scientists to uncover the true nature of this famous extinct bird. But we are not just looking back in time - our research could help save today's endangered birds too.
Using cutting-edge computer technology, we are piecing together how the Dodo lived and moved. This isn't just about satisfying our curiosity. By understanding how birds evolved in the past, we are learning valuable lessons that could help protect bird species today. It's like solving a 300-year-old puzzle, and the solution might just help us prevent more birds from going the way of the Dodo.
Professor Marcus O Heller, co-author
Bioengineering Research Group
Faculty of Engineering and Physical Sciences
University of Southampton, Southampton, UK.
Palaeoartist Karen Fawcett’s Dodo sculpture
The project will include work with palaeoartist Karen Fawcett , who has created a detailed, life-size model of the Dodo to bring the words on the pages of books and journal articles to life.
This work has been the merging of science and art to achieve accuracy and realism so that these creatures come back from the dead, real and tangible for people to touch and see.
Karen Fawcett
Palaeoartist
The work is supported by the University of Southampton’s Institute for Life Sciences.
The Institute was delighted to support this exciting work which exemplifies Southampton’s strength in interdisciplinary research and advanced scholarship.
Professor Max Crispin.
Director
Institute for Life Sciences.
University of Southampton, Southampton, UK.
Creationists might want to ignore or prepare to lie about the following, especially the section on the terminology and nomenclature background, which contains a description of how taxonomy has adapted to the modern synthesis of evolutionary theory, cladistics, shared common ancestry and monophyletics as it will make distressing reading for those who have been fooled into believing that biologists are abandoning the TOE as not fit for purpose, since nothing could be further from the truth, as this paper shows:
Abstract
The Dodo and its extinct sister species, the Solitaire, are iconic exemplars of the destructive capabilities of humanity. These secondarily terrestrial columbids became extinct within a century of their first encounter with humanity. Their rapid extinction, with little material retained in natural history collections, led 18th and some early 19th century naturalists to believe that these aberrant birds were mythological. This meant that the nomenclatural publications in which their scientific nomina were established were based on accounts written before the species became extinct. As such, no type specimens were designated for either the Dodo or the Solitaire. Our in-depth historical overview of both species and associated family-group nomina found that the nominal authority of the Dodo-based family group is not what is reported in the literature. Moreover, our detailed review of the family-group nomina based on columbid genera ensures that the current columbid family-group systematization is valid. Changing nomenclatural norms between the 19th and 20th centuries had a profound impact on Dodo nomenclature; so much so that the Dodo is an example of how pervasive nomenclatural ‘ripples’ can be and a warning for our current world of multiple nomenclatural codes.
INTRODUCTION
The Mauritian Dodo, †Raphus cucullatus (Linnaeus, 1758) (Fig. 1), and the Rodrigues Solitaire, †Pezophaps solitaria (Gmelin, 1789) (Fig. 2), are textbook examples of evolutionary transitions and of human-made extinctions. Their morphologies were so aberrant that for a time, during the 18th and early 19th centuries, they were considered mythological (Duncan 1828, de Blainville 1835, Strickland 1844, 1848, Hume 2006; see Figs 1, 2). As said by Strickland (1848: 4): ‘So rapid and so complete was their extinction that the vague descriptions given of them [Dodo and Solitaire] by early navigators were long regarded as fabulous or exaggerated, and these birds, almost contemporaries of our great-grandfathers, became associated in the minds of many persons with the Griffin and the Phœnix of mythological antiquity’. The existence of the Solitaire, in particular, was long doubted, because for several decades it was known solely from the descriptions by Leguat (1708). Strickland (1844: 324) mentioned that the Solitaire had been considered either ‘fictitious, or to be founded on an imperfect description of the true Dodo’.
Dodo (†Raphus cucullatus) mounted composite skeleton [NHMUK S/1988.50.1 (PV A 3302)]. A, cranial view. B, left lateral view.
Solitaire (†Pezophaps solitaria) mounted skeletons (on display at the Royal College of Surgeons, London, UK in 2023). A, female individual. B, male individual. Note the difference in skeleton size and robusticity between the sexes.
A series of key papers during the early 19th century ‘resurrected’ the Dodo and the Solitaire from the realm of the mythological to the material (Duncan 1828, de Blainville 1835, Strickland 1844). The seminal work of Strickland (1848) and Melville (1848.1), in their shared volume, described in detail the anatomy of specimens still found in European collections at that time, in addition to giving an authoritative account of the history of the two species. However, it was not until new expeditions to the islands of Mauritius and Rodrigues during the 1860s that new incomplete skeletons of both species were discovered. The skeletal remains discovered in the ‘Mare aux Songes’ marsh during 1865 (Clark 1866, Hume et al. 2009) allowed the Dodo to be described more fully (Owen 1866.1), and the Solitaire was described by Newton and Newton (1868, 1869) after the Jenner excavations of 1865 discovered skeletal remains (Parish 2013: 234; Hume et al. 2015).
There has been renewed interest in the biology of the Dodo and the Solitaire in the 21st century. Studies have explored Dodo body mass (Brassey et al. 2016, van Heteren et al. 2017) and bone histology (Angst et al. 2017.1), and the endocranial anatomy of both species has been reconstructed digitally from computed tomography scans (Gold et al. 2016.1). New Dodo material has been discovered from Mare aux Songes, and the ecosystem of the Mare aux Songes Lagerstätte has been studied (see Rijsdijk et al. 2009.1, 2015.1, Meijer et al. 2012). The remarkable ‘Thirioux Dodos’ have been described in-depth, which includes the most complete Dodo skeleton known (Claessens and Hume 2015.2; Claessens et al. 2015.3). There have even been attempts to reconstruct digitally how these animals would have looked (Rodríguez-Pontes 2016.2). With each decade, our understanding of these aberrant birds is being revolutionized. To ensure that this work is on a firm basis, we need to ensure that the alpha and beta taxonomy (and accompanying nomenclature) of both species is stable. As we will show, there are no known type specimens for either species. Moreover, given that the use of Dodo-based (i.e. †Raphus) family-group nomina is now accepted within columbid systematics, we need to ensure that these names are themselves valid, in order to maintain the nomenclatural stability of extant pigeons and doves. To those ends, we provide an in-depth historical overview of the Dodo, the Solitaire, and the family-group nomina based upon them. We also establish a new nomen to unite both species: †Raphina.
Terminology and nomenclatural background
Before starting our historical overview, it is worth stating that the current rules of zoological nomenclatural are ‘relatively’ recent and have evolved from prior rules/suggestions made during the 19th century. We wish this to be clear from the outset, in order that readers will not mistake our comments hereafter as undue criticisms of past workers. There have also been dramatic shifts in both systematics (the paradigms and methods used to hypothesize clades) and nomenclature (the establishment of names for said clades, and the rules governing those names) between the 18th and 21st centuries. During the 18th and 19th centuries, the rules and norms of zoological nomenclature were being developed (e.g. Linnaeus 1758, Kirby 1815, Westwood 1836, 1837a, 1837.1b, Strickland 1837.2, 1878, Strickland et al. 1843, Dall 1878.1, Société Zoologique de France 1881, Douvillé 1882, American Ornithologists’ Union 1886, Blanchard 1889, Bütschli et al. 1893), prior to their widespread formulation and promulgation during the 20th century (ICZN 1905, 1961, 1964, 1985, 1999). Moreover, the paradigms used to hypothesize taxa were distinctly different, with the transition from a pre-evolutionary paradigm to an acceptance of paraphyletic groupings and groups united based on shared similarity, which then shifted to our current paradigm based on shared common ancestry and monophyletic groups (for a general overview of thought, see Mayr 1942, 1965, 1982, Hennig 1966, Nelson 1973, de Queiroz 1988, Mishler 2009.2; and for some clade-specific examples, see Allard et al. 1999.1, Dornburg and Near 2021, Cotterill et al. 2014 and the references therein).
The current International Code of Zoological Nomenclature (the Fourth Edition, ICZN 1999, 2003, 2012.1, 2016.3; ‘Zoological Code’ hereafter) is a direct descendent of ‘Blanchard’s Code’ (Blanchard 1889) via the Règles Internationales de la Nomenclature Zoologique [International Rules of Zoological Nomenclature] (ICZN 1905). Raphaël Blanchard, the ‘father of International Zoological Nomenclature’ (Bock 1994: 33), was the Chair of the nomenclatural committee of the International Congress of Zoology, the first President of the International Committee on Zoological Nomenclature, and the Editor of the French edition of the Règles Internationales. For the first International Congress of Zoology, he wrote an overview of zoological nomenclature and outlined what he believed would be an acceptable set of rules for the international corpus of zoologists (‘Blanchard’s Code’; Blanchard 1889). ‘Blanchard’s Code’ did not exist in a vacuum, because a plethora of nomenclatural codes for zoology had been proposed during the 19th century, with the earliest comprehensive code being proposed by the British Association for the Advancement of Science (‘Strickland’s Code’).
‘Strickland’s Code’ (Strickland et al. 1843) was formulated by a committee of British zoologists and palaeontologists (including famous individuals, such as Charles Darwin and Richard Owen, in addition to Hugh Strickland, who was pivotal in our understanding of the Dodo and the Solitaire), who set down many of the norms we recognize today; norms of the so-called ‘Linnean’ system of nomenclature, although this is perhaps more accurately called ‘Linnean–Westwoodian–Stricklandian’ nomenclature (sensuDubois 2011: 4–5). However, there were some important differences between ‘Strickland’s Code’ and the current Zoological Code (ICZN 1999), such as the proposed ‘start date’ for zoological nomenclature, which in ‘Strickland’s Code’ was 1766, beginning with the publication of the 12th edition of Systema Naturæ (Linnaeus 1766). The ensuing controversy over the ‘start date’ for zoological nomenclature cost ‘Strickland’s Code’ support amongst zoologists (Linsley and Usinger 1959: 41), with Dall (1878.1: 15) noting that the starting point used by the British Association had begun ‘admitting to recognition some ichthyological works printed between the dates of the tenth and twelfth editions [of Systema Naturæ]’. Other national societies began proposing their own nomenclatural codes, including the American Association for the Advancement of Science (Dall 1878.1), the Société Zoologique de France (Société Zoologique de France 1881), the American Ornithologists’ Union (American Ornithologists’ Union 1886), and the Deutsche Zoologische Gesellschaft (Bütschli et al. 1893), as did the Congrès international de géologie [International Congress of Geology] (Douvillé 1882). It was ‘Blanchard’s Code’ (Blanchard 1889) and the subsequent Règles Internationales (ICZN 1905) that would begin to bring international stability to zoological nomenclature (for further details, see Linsley and Usinger 1959, Bock 1994).
Zoological nomenclature of the 18th and early 19th centuries did not adhere to the quasi-legal system in place today. The renaming of pre-existing genera and specific epithets was commonplace (particularly up to the 1840s–1850s). Therefore, readers should not be surprised that the principal of priority with regard to nominal authority was not adhered to in Dodo nomenclature during this time period or that the formulation of names does not meet the requirements on the Zoological Code as we understand it today (ICZN 1999). It is also worth noting that when the Dodo and Solitaire were first named (Linnaeus 1766, Gmelin 1789), the concept of nomenclatural types did not exist. Witteveen (2016.4: 156) credited Westwood (1837a) as the originator of this concept, which then became incorporated into ‘Strickland’s Code’ (and subsequent nomenclatural codes). As such, type specimens were not designated for the Dodo or the Solitaire.
Before continuing, we also need to define the terminology we will be using. We will follow the suggestions and recommendations of Dubois and Fitzhugh. Dubois (2021.1: 39) noted that, ‘the term taxonomy is traditionally used in two distinct senses, to designate either a scientific discipline, or any scientific classification of organisms produced by this discipline and adopted as valid by taxonomists’. In order to distinguish between both meanings, Dubois (2005: 406) erected the term ergotaxonomy for the latter (‘classification used by a given author in a given work’, Dubois 2006.1: 250). To remove any ambiguity, we use the term ergotaxonomy to refer to any ‘taxonomic framework’ considered valid by their proposer.
We will use the term ‘systematics’ rather than ‘taxonomy’ throughout. There is disagreement within the field of evolutionary biology regarding whether taxonomy and systematics are different subfields (e.g. Simpson 1961.1, Wiley and Lieberman 2011.1), whether taxonomy is a subdiscipline within systematics (e.g. Michener et al. 1970, Dubois 2006.1, Pavlinov 2013.1, Winsor 2023 and the references therein), or whether systematics is a subfield of taxonomy (e.g. Toepfer 2011.2). However, others, such as Mayr and Ashlock (1991) and Fitzhugh (2008), have proposed that taxonomy is a synonym of systematics. We will follow Fitzhugh (2008: 54) and use the term ‘systematics’ throughout.
We also use the term ‘systematization’ in preference to ‘classification’ following Fitzhugh (2008). Fitzhugh (2008: 54) defined classification as the ‘segregation of objects into classes based on specified properties’, whereas systematization is ‘the organization of observations into a system of concepts, in the form of hypotheses, according to theory’ (the definitions of these terms given by de Queiroz 1988: 241 was similar). We consider the latter to be the best description of systematics, because both species and ‘higher-level’ clades are explanatory hypotheses rather than objects (e.g. see Fitzhugh 2005.1, 2008, Mortimer et al. 2021.2).
Herein, we follow ornithological convention and capitalize English vernacular names of species (Parkes 1978; and the International Ornithological Committee World Bird List v.13.2; https://www.worldbirdnames.org/english-names/spelling-rules/). Moreover, we use the English vernacular names for columbid species that appear in the International Ornithological Committee World Bird List v.13.2 (https://www.worldbirdnames.org/new/bow/pigeons/), but with the following exceptions: (i) Didunculus strigirostris (Jardine, 1845) is referred to as the Samoan Tooth-billed Pigeon, because another species (†Didunculus placopedetesSteadman, 2006.2) was present throughout the islands that constitute the Kingdom of Tonga until ~2850 years ago (Steadman 2006.2; Worthy and Burley 2020) and was also present on Efate Island, Vanuatu (Worthy et al. 2015.4); and (ii) we generally refer to †Pezophaps solitaria as the Solitaire rather than the Rodrigues Solitaire, in order that it is consistent with the use of ‘the Dodo’ for †Raphus cucullatus (i.e. not using Mauritian Dodo). We follow Dubois (2000: 39) in using the term nomen (plural nomina) for any ‘scientific name’ that is formulated in compliance with a nomenclatural code, which, in this case, is the Zoological Code (ICZN 1999, 2003, 2012.2, 2016.5).
Our open nomenclature and synonymy lists follow the recommendations of Richter (1948) (see: Matthews 1973.1, who outlined them in English, and Becker 2001, who gave a recent overview in German), Sigovini et al. (2016.6), and Horton et al. (2021.3). Finally, we use the dagger (†) symbol in front of nomenclatural nomina that denote extinct taxa (except when they appear in quotations).
The dodo was named before biologists understood about evolution and believed species were created without ancestors, so the idea of a systematic nomenclature which reflected the evolutionary relationships between species had not occurred to them, so the dodo and its close relative, the Rodriguez Island Solitaire, were given different family names. This new study proposes a new family name, Raphina, for both species which reflects their evolutionary relationship and puts them in their correct position in the evolutionary tree of the pigeons and doves (the columbids) based on a DNA analysis.
The fact that human agency could drive a species to extinction was a shock to the early biologists who subscribed to the 'creation' mythos and assumed that what God had created mankind would be unable to destroy. We now know that both parts of the superstition were wrong and disastrously so. Not only are species the product of their environment but can be destroyed simply by changing that environment. There is no god playing any part in the process and least of all protecting its 'creation'.
This realisation made the Oxford dodo an object of fascination especially for academics like Do-Do-Dodgson who saw in it a metaphor for revealed wisdom about the nature of reality and the difference between it and human expectations, based on religious mythology.
Creationists might find it distressing to read how modern taxonomy has been revised to reflect the modern evolutionary synthesis, recognising that species can be arranged in clades and family trees based on how they evolved and diversified from common ancestors which themselves were the result of diversification from earlier common ancestors. They will probably find it distressing too how a DNA analysis of an extinct species like the dodo is confirming that view of the origins of biodiversity and no sign of that impending abandonment of the Theory of Evolution that their cult leaders have been assuring them is about to happen, any day now, real soon.
Evolution, or more precisely change in allele frequency over time, inevitably records selection pressures on a species resulting in a genome which, when correctly read and compared to predecessors, should tell the story of changes in the species environment.
This principle is illustrated by the threatened species of bumblebee, the rusty patch bumblebee, Bombus affinis, which has recently declined by about 90% in the USA and is now considered an endangered species. If this rate of decline continues the species will probably be extinct within 20 years. This level of intense selection pressure has inevitably left its mark on the genome of the species.
On of the problems facing the species is the result of the way bees breed. The queen can normally produce two sorts of egg - a fertilised, diploid egg which will develop into a female and a haploid, unfertilised egg which will develop into a male or drone. The problem arises when there is a high level of inbreeding, due, for example, to a small population - which the bee is now facing. In that situation, the female can produce diploid males because both sets of chromosomes can be identical. Diploid males are normally sterile so reducing the breeding success of the local species.
One of the great crimes of the Abrahamic religions is the allegedly 'God-given' dominion over the entire planet Earth, its wildlife, its mineral wealth and its land and sea to humankind, to be treated as free and there for the sole benefit of humankind with no other purpose.
In the British Isles this has resulted in a landscape dominated by towns and cities, agriculture and monoclonal forestry, and coastal waters where anything edible is hoovered up and consumed, leaving, in many cases, stocks too small and immature, or too scarce to maintain a stable population, let alone recover.
And our waste in the form of single-use plastics, sewerage, industrial waste such as CO2 and heavy metals, agricultural run-off containing artificial fertilisers, have polluted and destroyed many waterways. Agricultural monocultures have produced virtual deserts, so far as many species are concerned and destroyed soil structure with over-use of pesticides and artificial fertilisers has led, in a few years, to loss of precious topsoil that took hundreds of thousands of years to create and ploughed-up water-meadows have gone, taking their biodiversity with them.
Consequently, our wild bird, wild mammal, insect and wild plant populations have been in steep and accelerating decline for most of the last two centuries.
According to the following report, we are now faced with the extinction of 1,500 of our 10,000 species. The report, "State of Nature" is the result of a collaboration between a large number of British and Irish conservation and wildlife charities. It can be read here:
The State of Nature report referred to in the article may be read here:
In southern Ontario, Canada, in 1866, a flock of migrating passenger pigeons, was estimated to be 1.5 Km (.93 miles) wide and 500 Km (310 miles) long. It took 14 hours to pass and contained an estimated 3.5 billion birds. The passenger pigeon was then probably the most abundant species of bird on Earth and certainly in North America.
On September 1, 1914, at Cincinnati Zoo, less than 50 years later, 'Martha' the last known passenger pigeon died and the species became extinct. The species had been functionally extinct when the last male died some years earlier. Its death went unrecorded. The last wild passenger pigeon is believed to have been shot in 1901.
The extinction of this species was due entirely to human intervention, including hunting for cheap meat on a massive scale, deforestation and habitat destruction. Because of its habit of migrating in closely-packed flocks, a single shot could bring down several birds.
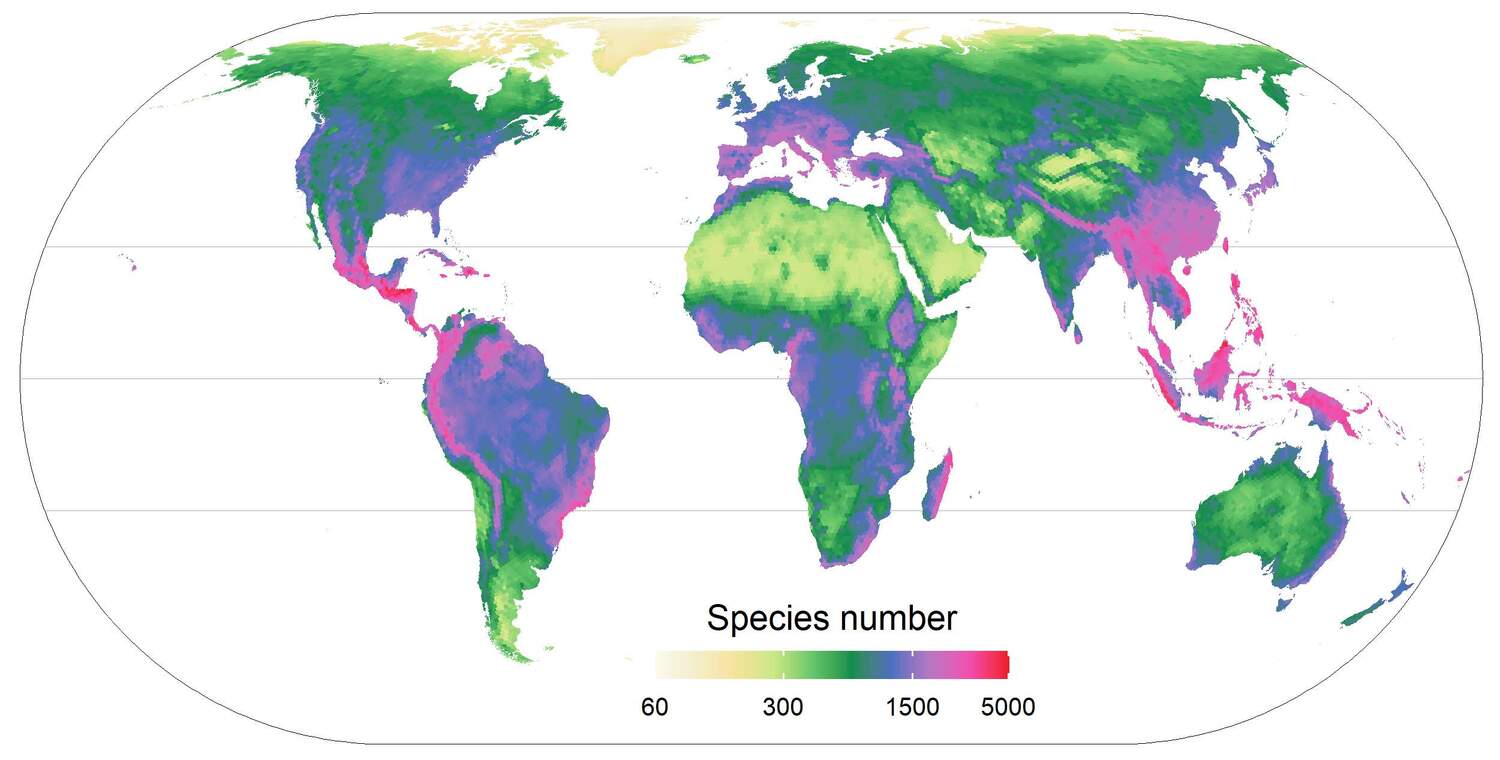
Global distribution of plant species richness across the globe as predicted using the distribution 300,000 plant species across 830 regional floras worldwide.
Elephants in Gorongosa National Park, Mozambique, declined by 90% during
the 20-year civil war because of ivory poaching. This spurred the rapid
evolution of tuskless females, such as this female
Loxodonta africana.
Since evolution by natural selection is driven primarily by the selecting
environment, rapid change in the environment can produce rapid evolution. If the
selection pressure is strong enough, this intense selection can also result in a
linked deleterious gene increasing in the population by being dragged up the
fitness landscape by a strongly advantageous gene to which it is linked by close
proximity on a chromosome.
This was the case in Mozambique, where during a 15-year-long civil war when
there was intense poaching of elephants for their ivory to finance the armies of
both sides and because any wild-life protection was largely absent or
non-functional. In the Gorongosa National Park for example, the elephant
population fell by 90% in 20 years with predation heavily selecting against
those with tusks. Consequently, the gene for tusklessness in females increased
in the population. Over a period of 28 years which included the 15 years of
civil war, the proportion of tuskless females increased from 18.5% (n = 52) to
50.9% (n = 108), but so did the proportion of females in the population and the
pre-term deaths of male calves!
This was due to an increase in the genes AMELX and MEP1a, which are associated
in mammals with normal tooth formation. AMELX is linked by its proximity on the
X chromosome to a dominant male-lethal syndrome in humans that also diminishes
the growth of maxillary lateral incisors (homologous to elephant tusks). In
other words, as AMELX increases in the population, so does a male-lethal gene
that also reduces the growth of tusks in elephants. The result is an absolute
increase in the proportion of live births of females and of tuskless females in
particular.
In the abstract to their recent paper, a team led by Princeton scientists Shane
C. Campbell-Staton of the Department of Ecology and Evolutionary Biology and
Brian J. Arnold of Department of Computer Science, Princeton University,
Princeton, NJ 08540, USA. said:
Abstract
Understanding the evolutionary consequences of wildlife exploitation is
increasingly important as harvesting becomes more efficient. We examined the
impacts of ivory poaching during the Mozambican Civil War (1977 to 1992) on
the evolution of African savanna elephants (Loxodonta africana) in
Gorongosa National Park. Poaching resulted in strong selection that favored
tusklessness amid a rapid population decline. Survey data revealed
tusk-inheritance patterns consistent with an X chromosome–linked dominant,
male-lethal trait. Whole-genome scans implicated two candidate genes with
known roles in mammalian tooth development (AMELX and MEP1a), including the
formation of enamel, dentin, cementum, and the periodontium. One of these loci
(AMELX) is associated with an X-linked dominant, male-lethal syndrome in
humans that diminishes the growth of maxillary lateral incisors (homologous to
elephant tusks). This study provides evidence for rapid, poaching-mediated
selection for the loss of a prominent anatomical trait in a keystone species.
Over the short term, this attrition of males is not overly detrimental to
elephants because herds are matriarchal with a single male able to serve
multiple females and 'spare' males forming non-breeding 'batchelor' herds, so
the population can remain stable or even increase with relatively few males.
What we can observe in this population then is a rapid evolution of a strong
advantageous gene, in that tusklessness makes females less likely to be poached,
with the 'opportunistic' increase in a deleterious gene which kills pre-term
male calves by being linked physically to a strongly advantageous one, the
deleterious effect in the population being more than compensated for by the
overall advantageous effect.
Over the longer term, and with control of poaching, whatever environmental
pressures led to the evolution of tusks in females originally is likely to
reassert itself and the balance should shift back towards tusked females and
more male live births. An beautiful example of how a changing environment
changes the meaning of the same genetic information without any change in that
information itself. And no magic deities involved anywhere in that process.
Lateral detail of a living Trioceros wolfgangboehmeisp. nov. from Goba, Ethiopia, showing the heterogeneous body scalation with both small scattered tubercles and enlarged flattened plate-like scales. In this individual the dorsolateral stripe is interrupted and forms a Y-shaped pattern on the flanks.
Photo by Petr Nečas.
7 / 8
Living juvenile of Trioceros wolfgangboehmeisp. nov. from Goba, Ethiopia.
Photo by Petr Nečas.
8 / 8
Digital elevation map of Ethiopia (generated by using the geographic information system ArcGIS 10.0; elevation in m a.s.l) indicating the currently known distribution of Trioceros wolfgangboehmeisp. nov. east of the Ethiopian Rift in the northern Bale Mountains (red stars; left star: Dinsho, right star: Goba). Black star: Addis Abeba. Grey dots show records of T. affinis based on distributional data after Largen and Spawls (2010) and Ceccarelli et al. (2014).
Here is another example of why fragile and vulnerable habitats need to be
preserved, if only for the rich variety of new species that could be living
there.
This time it is a new species of chameleon from the Bale Mountain forests of
Ethiopia. This one was discovered by zoologists Thore Koppetsch and Benjamin
Wipfler of the Research Museum Alexander Koenig in Bonn, Germany, and Petr Nečas
from the Czech Republic. The new species, Trioceros wolfgangboehmei is a
new small-sized chameleon living on the edge of the forest. Their findings were
published in the open-access, peer-reviewed life science journal
Zoosystematics and Evolution.

























{kind=link}