Had the ancient inhabitants of the Iberian Peninsula contrived to get their oral history and origin myths written down in the same way that the Bronze Age Canaanite pastoralists did, we might have had a slightly more accurate and less implausible history. It certainly wouldn't have had daft tales about a global genocidal flood just at the time newcomers with new ideas, a new language and new techniques of animal husbandry were migrating into the area having traversed Western Europe from the Steppes of Central Asia.
Unlike the parochial Canaanite nonsense, it might well have had people, places and animals from more than a day or two's walk from the Canaanite Hills.
Archaeological evidence shows that, at the time when creationist superstition says everyone had been drowned in a genocidal, global flood several thousand feet deep, by a vindictive god, people originating on the Steppes of Central Asia were migrating across western Europe and into the Iberian Peninsula, as though nothing unusual was happening.
In the study, scientists performed a genetic analysis on the bones of 133 human individuals from late Neolithic megalithic graves near Warburg in North Rhine-Westphalia. The team discovered the genome of the bacterium Yersinia pestis in the bone samples of two independent individuals. Additionally, previously published genomic data from a bone sample of a Neolithic dog found in Ajvide (Sweden) suggested a potential infection route.
What was creationism's divine malevolence up to with one of its most successful pathogens with which if killed hundreds of millions and changes society - the Yersinia pestis bacterium which caused the waves of black death and plague that regularly spread across the world?
It seems to have been experimenting, possibly trying to either perfect its virulence or work out the best delivery system to ensure it got to and killed as many people as possible. Sometimes, entire villages were wiped out. Not far from where I currently live are a couple of former villages that disappeared during the black death - the village of Woodperry near Oxford is an example, surviving now only in the name 'Woodperry Road' and a farmhouse later built on the site.
But 5000 years ago, Y. pestis doesn't seem to have been anything like a virulent as it became in the 12th Century. According to a recent discovery, it was capable of killing the occasional neolithic farmer but not of becoming a major pandemic able to kill hundreds of thousands and depopulate vast areas.
So, what changed, and more to the point, which explanation would a creationist prefer; the one which blames their god or the one which attributes it to evolution, climate change and cultural changes in human society? One thing we can be sure of though is the Michael J. Behe's biologically nonsensical religious apologetic of 'genetic entropy', causing the bacterium to 'devolve' away from an assumed created perfection (as though that were remotely possible), can be ruled out, because whatever the changes were, it led to a massive increase in the number of Y. pestis organisms, so was indisputably beneficial to it - in other words, in classical terms, it evolved.
The basis for any evolutionary change is environmental change. Providing that change is slow enough for living organisms to adapt they will adapt and change as the environment changes. This is how life contrives to make Earth look as though it is fine-tuned for life when the reality is that life is fine-tuned for Earth and the tuning mechanism is evolution.
But ask any small-town, American Bible Belt creationist and they will assure you Earth was perfectly designed for life, especially life of the human variety. It's pretty much like small-town American Bible Belt country, all over!
In fact, throughout its history, 99.9975% of which occurred before creationists think Earth was created, Earth has been in a state of dynamic change which, as soon as life manages to get close to tuning itself to it, it’s changed again. The underlying cause of this is mostly plate tectonics which keeps on breaking landmasses apart, sending them scudding across the surface of the planet from the tropics to the poles and ramming them into one another again, creating uplift mountains, deep ocean trenches and volcanos which keep pumping out greenhouse gasses and changing the climate, sometimes quite suddenly.
To illustrate this, a team of geologists, led by Xianzhi Cao of Ocean University of China, and including Professor Allan S. Collins of The University of Adelaide, Australia have reconstructed the last 1.8 billion years of Earth's geography.
How they did it and what it tells us is the subject of an article by Professor Collins in The Conversation. His article is reprinted here under a Creative Commons licence, reformatted for stylistic consistency:
In a very nice example of how humans and their gut microbiome have co-evolved, researchers from the Wellcome Sanger Institute, University College London (UCL), and the University of Birmingham, have identified a bacterium that helps digest breast milk and protects the baby from harmful pathogens.
As an example of intelligent [sic] design, this comes as close to a Heath-Robinson solution to a problem of the designer's own making as it's possible to imagine.
No designer who designed a baby's digestive system so it is susceptible to the harmful pathogens that it also designed, and created the baby to feed on breast milk that is difficult to digest, then solved those problems by creating another bacterium to fight the pathogens and help digest the milk, and infecting the baby with it, doesn't deserve the adjective 'intelligent'. 'Bloody stupid' would be a more appropriate term.
It's almost exactly as though the designer of these systems is a mindless automaton, simply obeying the natural laws of chemistry and physics and working without a plan!
The researchers have completed the largest study of UK baby microbiomes to date, carrying out whole genome sequencing to analyse stool samples from 1,288 healthy infants, all under one month old from the UK Baby Biome Study1.1.
Not content with the suffering that gum disease and dental caries cause, creationism's divine malevolence used at least one of the pathogens behind those problems to increase suffering a little more by aggrevating rhuematoid arthritis.
That's the inevitable conclusion honest creationist, who rejects the notion of evolution in which pathogenic parasites evolve naturally without intent, malevolent, benign or indifferent, should be drawing from the evidence from a Tokyo Medical and Dental University team of researchers led by Tokuju Okano and Toshihiko Suzuki of the Department of Bacterial Pathogenesis recently published in the International Journal of Oral Science.
This is not the first instance of creationism's divine malevolence multi-tasking its pathogens: in 2017 a team of researchers from The Chinese University of Hong Kong found a link between five oral bacteria, Peptostreptococcus stomatis, Streptococcus anginosus, Parvimonas micra, Slackia exigua and Dialister pneumosintes and the incidence of stomach and eosphageal cancer.
The oral bacterium which has been found to be aggrevating rheumatoid arthritis is Aggregatibacter actinomycetemcomitans.
One of the lines of attack against the science of evolutionary biology is the lie that Richard Dawkins was advocating selfishness with his seminal book, The Selfish Gene, so 'proving' that rejection of the Christian god as the explanation for biodiversity is because 'Evolutionists' just want to sin.
It is, of course, like other creationist attacks on science, utterly devoid of any factual basis and reflects badly both on those who seek to fool their target dupes with it and on their dupes who eagerly believe them in order to justify a pretense of moral superiority.
In fact, natural selection and differential success of different alleles within a selective environment is devoid of any moral contents because it is devoid of intelligent input and genes are passive in the process. In Dawkin’s analogy, the result is as though genes behave selfishly, not that they make moral decisions or have the ability to choose which environmental selectors act on them.
One of the predictable outcomes of this gene-centred approach is that in a competition, the result which suits all competitors is cooperation since in a cooperative alliance, all the genes are winners, just as with a human group, a cooperative group is invariably more successful than a group of competing individuals in perpetual conflict.
Artist’s reconstruction of Badjcinus timfaulkneri scavenging on the carcass of Silvabestius michaelbirti, a wombat-like marsupial also from Hiatus Site.
88 years ago today, the last Tasmanian tiger died in Beaumaris Zoo, Hobart, Tasmania.
It was the last in a line of carnivorous marsupial going back at least 23-25 million years ago, in the extended period of Earth's pre-'Creation Week' history when creationists believe there was nothing out of which a magic man made of nothing made everything in 6 days, just 10,000 years ago. According to creationist mythology, the Tasmanian tiger would have had no ancestors, being magically made from dirt just like every other living species.
This makes it hard for creation to explain the evidence of all those fossils known to be far older than 10,000 years and showing evidence of the evolution of extant species from ancient ancestors as species diversified from a single common ancestor which lived hundreds of millions of years ago, but then facts were never a problem for counter-factual creationism.
What information do you have on the extinct Tasmanian tiger?
The Tasmanian tiger, also known as the thylacine (Thylacinus cynocephalus), was a carnivorous marsupial native to Tasmania, mainland Australia, and New Guinea. The thylacine looked somewhat like a medium-to-large dog with a slender body, a head resembling that of a wolf or dog, and distinctive dark stripes across its back and tail, giving it the nickname "Tasmanian tiger." However, it was more closely related to kangaroos and other marsupials.
Key Information about the Tasmanian Tiger:
Physical Description:
The thylacine was about the size of a large dog, with a length of up to 1.8 to 2 meters (including the tail) and a height at the shoulder of about 60 cm.
Its fur was short and coarse, typically tan or yellow-brown, with 13 to 20 dark stripes across its back and tail.
It had a long, stiff tail, similar to a kangaroo's, and could open its jaws incredibly wide, up to 120 degrees.
Habitat and Behavior:
The thylacine was a nocturnal and solitary animal, hunting at night and resting during the day in a den or hollow log.
It primarily fed on small to medium-sized animals, such as kangaroos, birds, and small mammals. Its diet may have also included carrion.
Extinction:
The thylacine was driven to extinction on the Australian mainland thousands of years ago, likely due to competition with humans and the dingo, a wild dog introduced to Australia.
In Tasmania, the species persisted until the 20th century. The last known wild thylacine was killed in 1930, and the last known individual in captivity died in 1936 at the Hobart Zoo in Tasmania.
Its extinction was primarily due to human activities, including hunting (encouraged by a bounty system), habitat destruction, and the introduction of diseases.
Conservation Efforts and "Rediscovery" Claims:
Despite being declared extinct, there have been numerous unconfirmed sightings and efforts to find surviving thylacines, but none have been scientifically verified.
Some scientific projects have explored the possibility of "de-extinction" using preserved DNA from museum specimens, but these efforts remain largely theoretical.
Cultural Significance:
The thylacine holds a unique place in Tasmanian culture and Australian folklore, often seen as a symbol of the tragedy of extinction and the need for wildlife conservation.
It appears on Tasmania's coat of arms and has been the subject of various books, documentaries, and studies.
Would you like to know more about any specific aspect of the Tasmanian tiger?
How the oldest ancestors of the Tasmanian tiger were discovered by palaeontologists from the University of New South Wales, Australia, is the subject of a Taylor & Francis news release:
Earliest known ancestors of the extinct Tasmanian Tiger discovered – and some could even eat the bones and teeth of their preyA marsupial which had an “extremely thick” jawbone, enabling it to consume even the bones and teeth of its prey is among a discovery of three, new ancient species of the modern Thylacines – otherwise known as the Tasmanian tiger, which went extinct 88 years ago.
These new species each roamed Australia around 23-to-25 million years ago, during the late Oligocene, making them the “undoubted oldest members of this family ever discovered”.
Today – Australia’s National Threatened Species Day, which marks the death of the last Tasmanian Tiger in Hobart’s Beaumaris Zoo on the 7th of September 1936 – scientists from the University of New South Wales (UNSW) Vertebrate Palaeontology Lab publish their findings in the Journal of Vertebrate Paleontology.
The once suggested idea that Australia was dominated by reptilian carnivores during these 25 million-year-long intervals is steadily being dismantled as the fossil record of marsupial carnivores, such as these new thylacinids, increases with each new discovery. The diversity of mammalian carnivores at Riversleigh during this period rivals that seen in any other ecosystem, including the great mammalian carnivore radiation that developed in South America.
Timothy Churchill, lead author
University of New South Wales
Sydney, New South Wales, Australia.
The three new species were each found in the fossil-rich deposits in Riversleigh World Heritage Area.
The largest of these new species, Badjcinus timfaulkneri, weighed somewhere between 7-11 kilograms, about the same size as a large Tasmanian Devil. Alike the Tasmanian Devil, timfaulkneri possessed an extremely thick jawbone enabling it to consume the bones and teeth of its prey. This species is related to the much smaller, previously discovered B. turnbulli (2.7 kg) – which until now was the only other undoubted thylacinid known from the late Oligocene.
The dentary and isolated first molar of B. timfaulkneri were recovered from Hiatus Site which is even older than Riversleigh’s White Hunter Site where B. turnbulli was previously found, making B. timfaulkneri the oldest undoubted thylacine discovered so far.
Badjcinus timfaulkneri is named after Tim Faulkner, the director and co-owner of the Australian Reptile Park and managing director of Aussie Ark. Tim has dedicated his life to the conservation of Australia’s wildlife including the largest still-living marsupial carnivore, the Tasmanian Devil.
The second new species is Nimbacinus peterbridgei. This was about the size of a Maltese Terrier (~3.7 kg). This species is represented by a near-complete dentary from White Hunter Site. Nimbacinus peterbridgei was a predator that probably focused on small mammals and other diverse prey species that lived with it in the ancient forests. Species of Nimbacinus appear to be more closely related to the Tasmanian Tiger than other thylacinids of similar age. This means Nimbacinus peterbridgei is probably the oldest direct ancestor of the Tasmanian Tiger yet known.
Nimbacinus peterbridgei was named after Australian geologist, speleologist and bibliophile Peter Bridge. He has devoted his life to helping uncover Australia’s ancient past, particularly in the caves of Western Australia.
The last species, Ngamalacinus nigelmarveni was a ~5.1kg thylacinid – approximately the size of a Red fox. It was also from White Hunter Site at Riversleigh. The blades on the lower molars of species of Ngamalacinus are elongated with deep V-shaped carnassial (‘meat-cutting’) notches, suggesting they were highly carnivorous – more so than any of the other thylacinids of similar size.
Ngamalacinus nigelmarveni is named after Nigel Marven, a renowned British television documentary presenter famous for paleontology-inspired series like Prehistoric Park and Sea Monsters.
The presence of three distinct lineages of specialised thylacinids during the late Oligocene highlights how quickly they diversified after first appearing in the fossil record. These thylacinids exhibits very different dental adaptations, suggesting there were several unique carnivorous niches available during this period. All but one of these lineages, the one that led to the modern Thylacine, became extinct around 8 million years ago.
That lineage of these creatures that survived for more than 25 million years ended with the death of Benjamin, the last Tasmanian Tiger in Hobart’s Beaumaris Zoo on the 7th of September 1936.
Professor Michael Archer, co-author
University of New South Wales
Sydney, New South Wales, Australia.
ABSTRACT
New thylacinid species of Badjcinus, Nimbacinus, and Ngamalacinus are described from upper Oligocene deposits of the Riversleigh World Heritage Area, northwestern Queensland. Badjcinus timfaulkneri, Nimbacinus peterbridgei, and Ngamalacinus nigelmarveni are among the oldest thylacinids yet known and indicate an earlier diversification of the family than previously understood. Maximum parsimony analysis supports a sister group relationship between Ng. nigelmarveni and Ng. timmulvaneyi, but the relationships of the two other new taxa are unresolved. Bayesian dated total evidence analysis using morphological and molecular data supports the generic assignment of B. timfaulkneri and Ng. nigelmarveni but not that of Ni. peterbridgei. Both phylogenies herein support a taxonomic reassignment of Thylacinus macknessi to the genus Wabulacinus, a conclusion also supported by the results of previous studies. Body mass estimates based on molar size regressions indicate body sizes ranging from 3.7 kg to 11.4 kg for the new thylacinid species. Badjcinus timfaulkneri exhibits an extremely deep jaw compared with other thylacinids, with mandibular bending strength analysis suggesting that it was a highly durophagous carnivore much like the modern dasyurid Sarcophilus harrisii. This analysis also suggests Ni. peterbridgei had a dentary more similar in shape to that of plesiomorphic thylacinid faunivores such as Ni. dicksoni and T. cynocephalus suggesting that it had a relatively more generalist faunivorous diet. The molars of Ng. nigelmarveni suggest they were better suited for longitudinal slicing than the molars of B. timfaulkneri and Ni. peterbridgei, indicating a more hypercarnivorous diet compared with that of those species.
INTRODUCTION
There are 12 extinct species in the dasyuromorphian family Thylacinidae, 10 of which are from the Oligo-Miocene (26–5.3 Ma) (Rovinsky et al., 2019). Unlike during the Plio-Pleistocene (5.3–0.12 Ma), which is dominated by large hypercarnivorous species of Thylacinus (15–55 kg), the Oligo-Miocene radiation exhibits considerably higher generic diversity, with seven monospecific clades known from the Riversleigh World Heritage Area in northwestern Queensland, all of which were between 3–10 kg in body mass. This restriction in size suggests thylacinids occupied most of the small- to medium-sized faunivorous niches at Riversleigh (1–10 kg). Contemporary peramelemorphians (∼50 g–1.5 kg) (Gurovich et al., 2014; Travouillon et al., 2013; Travouillon et al., 2010, 2014.1) and thylacoleonids (∼10–50 kg) (Gillespie, 2023; Gillespie et al., 2016, 2019.1a, 2019.2b) occupied relatively smaller and larger carnivore niches.
The oldest undoubted thylacinid, Badjcinus turnbulli Muirhead & Wroe, 1998, is known only from White Hunter Site in the Riversleigh World Heritage Area. This and other late Oligocene sites at Riversleigh have not yet been radiometrically dated. White Hunter Site is interpreted to be late Oligocene in age because of the presence of the ilariid Kuterintja ngama, otherwise only known from the Ngama Local Fauna from the Etadunna Formation, which is magnetostratigraphically dated to 24.8–25 Ma (Myers & Archer, 1997; Woodburne et al., 1994). Riversleigh’s Faunal Zone A (FZA) deposits contain taxa that support a late Oligocene age (Arena et al., 2016.1; Travouillon et al., 2006). The relatively plesiomorphic dentition of B. turnbulli has led to difficulty in taxonomic assignment, with phylogenetic analyses placing it either as a basally branching member of Thylacinidae (Kealy & Beck, 2017; Muirhead & Wroe, 1998; Murray & Megirian, 2006.1a; Wroe & Musser, 2001), as a stem dasyurid (Wroe et al., 2000), or as a sister group to Dasyuromorphia as a whole (Kealy & Beck, 2017).
Only two other thylacinid fossils are known from upper Oligocene deposits. Originally assigned to Nimbacinus dicksoni but later reassigned to Thylacinidae incertae sedis, an isolated m2 (QM F16809) from D-Site at Riversleigh is the only other thylacinid known from Riversleigh’s upper Oligocene deposits (Muirhead & Archer, 1989; Murray & Megirian, 2000.1; Wroe & Musser, 2001). An isolated, broken M2 (NTM P2815–10) of a thylacinid is also known from the Pwerte Marnte Marnte Local Fauna (LF) in the Northern Territory, a deposit assumed to be upper Oligocene on the basis of biocorrelation (Murray & Megirian, 2006.2b; Woodburne et al., 1994). Although this upper molar, which is similar in size to the putative thylacinid Mutpuracinus archibaldi, was suggested by Murray and Megirian (2006.2b) to be the oldest thylacinid in the fossil record, there are doubts about its identification as a thylacinid (see Discussion).
Nimbacinus dicksoni Muirhead & Archer, 1989, is the best preserved and researched Miocene thylacinid (Attard et al., 2014.2; Murray & Megirian, 2000.1; Wroe & Musser, 2001). Multiple specimens including a near complete skull and skeleton are known from Middle Miocene deposits at Riversleigh and from the Bullock Creek LF in the Northern Territory. The dentition of Ni. dicksoni is relatively more plesiomorphic than that of other thylacinids (except possibly Muribacinus gadiyuli) in retaining slightly reduced metaconids on m2–4 and unreduced stylar cusps on M1–3. Previous morphofunctional analyses of the skull of Ni. dicksoni suggest it was a voracious predator capable of hunting prey larger than itself, with biting capabilities most similar to extant species of Dasyurus, rather than to the larger Thylacinus cynocephalus (Attard et al., 2014.2).
In addition to Nimbacinus dicksoni, four monotypic thylacinid genera are known from upper and lower dentitions recovered from Early and Middle Miocene Riversleigh deposits. These include the medium-sized (∼5–7 kg) Wabulacinus ridei and Ngamalacinus timmulvaneyi Muirhead, 1997.1, as well as the diminutive (∼1–2 kg) Muribacinus gadiyuli Wroe, 1996, and the large (∼18 kg) Maximucinus muirheadae Wroe, 2001.1a (Myers, 2001.2). The dentition of Ng. timmulvaneyi, W. ridei, and Ma. muirheadae have been considered to be relatively plesiomorphic in comparison with species of Thylacinus, but more derived than Ni. dicksoni (Muirhead, 1997.1). The diminutive Mur. gadiyuli is dentally the most plesiomorphic thylacinid known (Wroe, 1996).
Thylacinus macknessi Muirhead, 1992, from Riversleigh’s Early Miocene Neville’s Garden Site, is tentatively regarded as the earliest known member of the genus Thylacinus (Muirhead & Gillespie, 1995). This taxon possesses unique dental adaptations associated with a shift within the Thylacinus lineage towards hypercarnivory, including near complete loss of metaconids on m2–4, loss of entoconids, reduction of the lingual portion of the talonid margin in m2–4 and a lingual shift of the hypoconid so that the cristid obliqua forms a continuous longitudinal blade with the paracristid.
The powerful thylacine, T. potens Woodburne, 1967, from the Alcoota Local Fauna in the Northern Territory, has been interpreted on the basis of biocorrelation to be Late Miocene between 8.5 and 5.5 Ma (Megirian et al., 1996.1, 2010.1). It is the largest and most hypercarnivorous thylacinid known. It has been estimated to be between 30–56 kg in adult body mass (Myers, 2001.2; Wroe, 2001.1a), with some estimates exceeding 120 kg (Yates, 2014.3). Two additional species of Thylacinus with dental adaptations for increased carnivory, T. yorkellus and T. megiriani, are known from Upper Miocene to Lower Pliocene deposits (Murray, 1997.2; Yates, 2015).
Tyarrpecinus rothi Murray & Megirian, 2000.1, is a small thylacinid from the Late Miocene Alcoota Local Fauna of the Northern Territory. It is known from a broken maxilla that retains P2 and an isolated M2. The phylogenetic relationships and paleobiology of this taxon cannot be confidently determined until more complete craniodental material is found.
The quoll-sized Mutpuracinus archibaldi Murray & Megirian, 2000.1, known from a near complete skull with an incomplete upper and lower dentition from the Middle Miocene Bullock Creek LF in the Northern Territory (Murray & Megirian, 2006.1a), was initially considered to be an early thylacinid because of craniodental and basicranial similarities to plesiomorphic thylacinids such as Ni. dicksoni. However, more recent phylogenetic analyses suggest it should be regarded as Dasyuromorphia incertae sedis because of its lack of craniodental synapomorphies uniting it with undoubted thylacinids (Churchill et al., 2023.1; Kealy & Beck, 2017; Rovinsky et al., 2019).
Two further medium-sized (1–10 kg) incertae sedis dasyuromorphians are known from Miocene deposits; Whollydooleya tomnpatrichorum Archer, Christmas et al., 2016.2, from Miocene deposits in New Riversleigh (an area approximately 10 km southwest of the Riversleigh World Heritage Area) and Apoktesis cuspis Campbell, 1976, from upper Oligocene deposits at Lake Ngapakaldi in the Tirari Desert of South Australia. Either may be related to thylacinids or dasyurids (Archer, Christmas et al., 2016.2; Campbell, 1976). However, the lack of adequate fossil material to critically assess the relationships of these two enigmatic taxa precludes a more precise assignment beyond Dasyuromorphia incertae sedis. Whollydooleya tomnpatrichorum is only known from a single lower molar while the location of the skull and dentaries attributed to A. cuspis is currently unknown.
In this study, we describe three new species of thylacinid from Riversleigh’s oldest deposits: the upper Oligocene Hiatus and White Hunter Sites. The fossil specimens herein are all dentaries retaining near complete or partial lower dentitions.
Of course, it would be stupid to assume the Bronze Age pastoralists who wrote the Bible knew anything about Australia, or even a southern hemisphere. Since they believe Earth was a small flat place with a dome over it, they wouldn't even have considered hemispheres, northern of southern. Their view of the world was so narrow and restricted they thought all they had to explain was the small area within a day or two's walk of the Canaanite Hills, so nothing outside that small area was included; not an animals, mountain, continent or people; nothing.
So, the idea that they would have believed the Tasmanian tiger was magically created without ancestors 10,000 years ago or less is utterly preposterous and could only be believed by someone at least as blind to the evidence as were the authors of Genesis.
If there is one thing that creationists get right it is that science is a threat to their childish superstition, so they have two main lines of argument: firstly they tell us how their religion disagrees with science, using Bible quotes as though they are indisputable facts, not just claims written down and declared to be true; secondly they attack science as though finding fault with one aspect or another of science somehow invalidates all of it and their superstition wins by default.
Neither of these tactics ever provide any evidence for creationism of course because there is none to produce, but they have the desired effect of making the creationist feel superior to those clever-dicky, elitist scientists with their big words.
And one of their favourite lines of attack is to claim that 'science can't explain consciousness', conveniently forgetting to add the word 'yet' to the end of their claim. A basic understanding of science and the history of science, would tell them that not yet understanding something doesn't invalidate science, it invalidates ignorance. Science never used to understand lightening or earthquakes, or diseases, or atoms, or biodiversity and the appearance of kinship between species, but now it does.
And now we are beginning to understand consciousness too. We know, for example, that it doesn't exist independently of a fully-functional brain because it can be abolished with chemicals and by injury, so we know that whatever the mechanism, it is a function of neurophysiology and so amenable to scientific investigation and explanation. The questions are the 'how' and the 'what' that makes up all scientific investigation.
Imaginative reconstruction of a European scene in the Aurignacian
AI generated by ChatGPT4.0
The distributions of density in (P 100 km−2) for a 43, b 42, c 41, d 40, e 39 and f 38 ka are shown. The symbols represent the AUR archaeological sites for different phases (full black dots: Phase 1; open circle: Phase 2; and red triangles: Core sites).
There are of course, very many things Bible-literalist creationists need to ignore, lie about and/or misrepresent to maintain the delusion of Earth being just a few thousand years old, not the least of which is the abundant geological and archaeological evidence of human activity long before they believe Earth was created by magic out of nothing in the so-called 'Creation Week'.
Their handicap is in trying to compress the entire 13.8 billion year history of the Universe, the 3.8 billion year history of planet Earth and the 2-3 million year history of human evolution, into 10,000 or fewer years, and then trying to ignore the evidence of continuous, unbroken cultural history extending from way before a global genocidal flood, right through it and continuing to modern times, as though such a flood never happened.
And, as is normal with science, a paper just published in Nature Communications, gives them more evidence to ignore, lie about and/or misrepresent. It shows the history of early modern human migration across Europe during the Aurignacian (43,000 -32,000 years ago). The research comes from a team from the University of Cologne, Germany, led by Professor Dr Yaping Shao. It is explained in a University of Cologne press release:
Most cells, either single-celled organisms like bacteria or the eukaryote cells of multicellular organisms, reproduce by simple division into two daughter cells. Under favourable conditions, this means a cell can produce a population of descendants that grows exponentially 1 → 2 → 4 → 8 → 16 → 32→64 … etc. (population = 2n; where n= the number of generations)
But the trick a bacterium, Corynebacterium matruchotii, that is only found in the human mouth uses is to divide into multiple new cells at each generation. For example, assuming it splits into 10 daughter cell at each generation, its growth rate from a single founder cell will be 1 → 10 → 100 → 1000 → 10,000 → 100,000 → 1,000,000 … etc., (population = 10n). But scientists have found it can do better than that, producing up to 14 new cells at each generation.
In 6 generations in favourable conditions, Corynebacterium matruchotii can produce 1 million or more offspring, against an 'ordinary' bacteria's 64, so rapidly out-pacing any other bacteria. C. matruchotii is an essential component of the plaque that quickly develops on teeth. It appears to have no other known function.
In some ways, C. matruchotii behaves more like fungal hyphae, growing at the tip to produce a long thin filament. It is this filament that then simply splits up into small sections, each of which becomes a new cell.
The filamentous bacteria C. matruchotii splitting into multiple cells at once, a rare kind of cell division called multiple fission.
Credit: Scott Chimileski, MBL
In 2020 a team of researchers showed how plaque is a highly structured and organised colony of different microorganisms which almost seems to be designed to allow Streptococcus mutans to get on with its job of dissolving the tooth enamel and causing caries. Creationists dogma insists that anything with a complex, organised structure must be intelligently designed because, so they assert, complexity and order can’t arise from disorder without intelligent intervention.
This latest discovery shows how the beginnings of this organised colony are established very quickly.
Not content with a whole range of parasitic worms to cause suffering, creationism's divine malevolence has also designed them to carry a payload of viruses to make their victims suffer even more.
This is the inescapable conclusion an honest creationist (if there is such a thing) would need to reach if he/she made an honest assessment of the facts just published by a team of researchers from the Centre for Neglected Tropical Diseases, Liverpool School of Tropical Medicine (LSTM), Liverpool, UK and the Global Health Institute, University of Antwerp, Antwerp, Belgium, in the journal Nature Microbiology.
This research team have identified a wide variety of viruses infecting nematodes and, most worryingly, the parasitic nematodes responsible for diseases in humans, so it is entirely possible that the diseases associated with infection by these nematodes could actually be caused by the viruses they carry. Related nematodes also show signs of virus-nematode co-evolution over millions of years in the form of a global distribution of nematode-virus associations.
Their research is explained in a press release from LSTM:
Tell me all about parasitic nematodes and the diseases they cause in humans, please.
Parasitic nematodes (roundworms) are a diverse group of organisms that infect humans, animals, and plants. They can be found in various environments, from soil to water, and have adapted to parasitize a wide range of hosts. These nematodes can cause various diseases in humans, ranging from mild discomfort to severe and life-threatening conditions.
Overview of Parasitic Nematodes
Parasitic nematodes are classified under the phylum Nematoda. They are elongated, cylindrical, and non-segmented worms. Some of the most common parasitic nematodes that infect humans include:
Ascaris lumbricoides (giant roundworm)
Trichuris trichiura (whipworm)
Ancylostoma duodenale and Necator americanus (hookworms)
Strongyloides stercoralis (threadworm)
Enterobius vermicularis (pinworm)
Wuchereria bancrofti and Brugia malayi (filarial worms)
Onchocerca volvulus (cause of river blindness)
Dracunculus medinensis (Guinea worm)
Toxocara spp (Toxocariasis)
Common Diseases Caused by Parasitic Nematodes in Humans
Ascariasis
Causative agent:Ascaris lumbricoides
Symptoms: Often asymptomatic, but heavy infections can cause abdominal pain, malnutrition, growth retardation in children, intestinal obstruction, and respiratory issues (due to larval migration through the lungs).
Transmission: Ingestion of eggs from contaminated soil, water, or food.
Geographical distribution: Common in tropical and subtropical areas with poor sanitation.
Trichuriasis (Whipworm Infection)
Causative agent:Trichuris trichiura
Symptoms: Asymptomatic in mild cases, but heavy infections can cause diarrhea, rectal prolapse, anemia, growth retardation in children, and malnutrition.
Transmission: Ingestion of eggs from contaminated soil, food, or water.
Geographical distribution: Widespread in tropical regions, especially in areas with poor sanitation.
Hookworm Infections
Causative agents:Ancylostoma duodenale and Necator americanus
Symptoms: Anemia (due to blood loss in the intestines), abdominal pain, diarrhea, weight loss, and fatigue. Skin itching and rash can occur where larvae penetrate the skin.
Transmission: Larvae penetrate the skin, usually through bare feet when walking on contaminated soil.
Geographical distribution: Common in warm, moist climates, particularly in sub-Saharan Africa, Southeast Asia, and South America.
Strongyloidiasis
Causative agent:Strongyloides stercoralis
Symptoms: Often asymptomatic, but can cause abdominal pain, diarrhea, skin rash, and severe life-threatening hyperinfection syndrome, especially in immunocompromised individuals.
Transmission: Larvae penetrate the skin from contact with contaminated soil.
Geographical distribution: Tropical and subtropical regions, but also occurs in temperate areas.
Enterobiasis (Pinworm Infection)
Causative agent:Enterobius vermicularis
Symptoms: Perianal itching, irritability, sleep disturbances, and sometimes abdominal pain.
Transmission: Ingestion of eggs from contaminated hands, surfaces, or food.
Geographical distribution: Worldwide, particularly in temperate regions; commonly affects children.
Symptoms: Lymphedema (swelling of limbs), elephantiasis (thickening of the skin and underlying tissues), and hydrocele (swelling of the scrotum).
Transmission: Mosquito bites (primarily Culex, Anopheles, and Aedes species).
Geographical distribution: Sub-Saharan Africa, Southeast Asia, the Pacific, and parts of the Americas.
Onchocerciasis (River Blindness)
Causative agent:Onchocerca volvulus
Symptoms: Severe itching, skin rash, depigmentation, and nodules under the skin. Can lead to visual impairment or blindness if the eyes are affected.
Transmission: Blackfly bites (genus Simulium).
Geographical distribution: Sub-Saharan Africa, with some cases in Latin America and Yemen.
Dracunculiasis (Guinea Worm Disease)
Causative agent:Dracunculus medinensis
Symptoms: Painful blister and ulcer, often on the lower limbs. The emerging worm can cause secondary infections.
Transmission: Ingestion of water contaminated with infected copepods (small crustaceans) that harbor larvae.
Geographical distribution: Historically widespread in Africa and Asia, now largely confined to a few countries due to eradication efforts.
Toxocariasis
Causative agents:Toxocara canis and Toxocara cati
Symptoms: In visceral larva migrans (VLM), symptoms may include fever, cough, abdominal pain, and eosinophilia; in ocular larva migrans (OLM), it may cause visual impairment or blindness.
Transmission: Ingestion of eggs from contaminated soil or food, or from contact with infected animal feces.
Geographical distribution: Worldwide, especially in areas where pets are not regularly dewormed.
Diagnosis and Treatment
Diagnosis of parasitic nematode infections typically involves stool examinations for eggs or larvae, blood tests for antibodies or microfilariae, or skin snips in the case of onchocerciasis. Advanced imaging or biopsies may be required for some infections.
Treatment often involves anti-parasitic medications such as albendazole, mebendazole, ivermectin, diethylcarbamazine, and praziquantel, depending on the type of nematode and the severity of the infection.
Prevention and Control
Improving sanitation: Proper disposal of human feces and access to clean water.
Personal hygiene: Handwashing, wearing shoes, and using protective clothing.
Mass drug administration (MDA): In endemic areas to reduce transmission, especially for lymphatic filariasis and onchocerciasis.
Vector control: Reducing the population of mosquitoes and blackflies.
Public health education: Raising awareness about transmission and prevention methods.
Would you like more information on any specific nematode or disease?
Newly discovered viruses in parasitic nematodes could change our understanding of how they cause diseaseNew research shows that parasitic nematodes, responsible for infecting more than a billion people globally, carry viruses that may solve the puzzle of why some cause serious diseases.
A study led by Liverpool School of Tropical Medicine (LSTM) used cutting-edge bioinformatic data mining techniques to identify 91 RNA viruses in 28 species of parasitic nematodes, representing 70% of those that infect people and animals. Often these are symptomless or not serious, but some can lead to severe, life-changing disease.
Nematode worms are the most abundant animals on the planet, prevalent in all continents worldwide, with several species infecting humans as well as agriculturally and economically important animals and crops. And yet in several cases, scientists do not know how some nematodes cause certain diseases.
The new research, published in Nature Microbiology(link is external)(opens in a new tab), opens the door to further study of whether these newly discovered viruses – only five of which were previously known to science – could contribute to many chronic, debilitating conditions. If a connection can be proven, it could pave the way for more effective treatments in the future.
This is a truly exciting discovery and could change our understanding of the millions of infections caused by parasitic nematodes. Finding an RNA virus in any organism is significant, because these types of viruses are well-known agents of disease. When these worms that live inside of us release these viruses, they spread throughout the blood and tissues and provoke an immune response. This raises the question of whether any of the diseases that these parasites are responsible for could be driven by the virus rather than directly by the parasitic nematode.
Professor Mark J. Taylor, co-corresponding author
Professor of Parasitology
Centre for Neglected Tropical Diseases
Department of Tropical Disease Biology
Liverpool School of Tropical Medicine, Liverpool, UK.
Parasitic nematodes including hookworms and whipworms can cause severe abdominal problems and bloody diarrhoea, stunted development and anaemia. Infection with filarial worms can lead to disfiguring conditions such as lymphoedema or ‘elephantiasis’, and onchocerciasis, or ‘river blindness’, that leads to blindness and skin disease.
This is a truly exciting discovery and could change our understanding of the millions of infections caused by parasitic nematodes.
The study authors propose that these newly identified viruses may play a role in some of these conditions. For example, Onchocerciasis-Associated Epilepsy (OAE) that occurs in children and adolescents in Sub-Saharan Africa has recently been associated with onchocerciasis, but it is not known why this causes neurological symptoms such as uncontrollable repeated head nodding, as well as severe stunting, delayed puberty and impaired mental health.
One of the viruses in the parasites that cause onchocerciasis identified in the new study is a rhabdovirus – the type that causes rabies. The authors of the study suggest that if this virus is infecting or damaging human nerve or brain tissue, that could explain the symptoms of OAE.
The full extent and diversity of the viruses living in parasitic nematodes, how they impact nematode biology and whether they act as drivers of disease in people and animals now requires further study.
The illuminating discovery of these widespread yet previously hidden viruses was first made by Dr Shannon Quek, a Postdoctoral Research Associate at LSTM and lead author of the new study, who had initially been using the same data mining method to screen for viruses within mosquitoes that spread disease, before deciding to investigate nematodes.
As a child [in Indonesian], I saw a lot of people infected with these diseases and I suffered from the dengue virus on three occasions. That got me interested in tropical diseases. Diseases caused by parasitic nematodes are very long-term, life-long illnesses that persistently affect people. It has a significant impact on people's quality of life, their economic outputs and mental health.
There are a lot of studies about the microbiomes of mosquitoes, and how the bacteria that lives inside can block the spread of viruses, which might stop vector-borne diseases like dengue. This interplay between organisms in the same host led me to think - what else might be inside parasitic nematodes as well? Which after my discovery will now be the focus of our research.
Dr Shannon Quek, lead author
Centre for Neglected Tropical Diseases
Department of Tropical Disease Biology
Liverpool School of Tropical Medicine, Liverpool, UK.
The study also involved researchers from University of Antwerp and KU Leuven, Belgium, Brock University, Canada, University of Queensland, Australia, University of Buea, Cameroon and the University of Energy and Natural Resources, Ghana.
Abstract
Parasitic nematodes have an intimate, chronic and lifelong exposure to vertebrate tissues. Here we mined 41 published parasitic nematode transcriptomes from vertebrate hosts and identified 91 RNA viruses across 13 virus orders from 24 families in ~70% (28 out of 41) of parasitic nematode species, which include only 5 previously reported viruses. We observe widespread distribution of virus–nematode associations across multiple continents, suggesting an ancestral acquisition event and host–virus co-evolution. Characterization of viruses of Brugia malayi (BMRV1) and Onchocerca volvulus (OVRV1) shows that these viruses are abundant in reproductive tissues of adult parasites. Importantly, the presence of BMRV1 RNA in B. malayi parasites mounts an RNA interference response against BMRV1 suggesting active viral replication. Finally, BMRV1 and OVRV1 were found to elicit antibody responses in serum samples from infected jirds and infected or exposed humans, indicating direct exposure to the immune system.
Main
Humans and animals are frequently infected with multiple species of parasitic nematodes1,2,3 and suffer from chronic, lifelong infections and exposure to continuous reinfection4. Such infections impose a substantial health burden on billions of people, impacting their health, quality of life and economic productivity. Medically important parasitic nematodes infect over one billion people, resulting in up to 7.53 million disability-adjusted life years globally5. Prominent examples include intestinal species such as Ascaris lumbricoides and Trichuris trichiura4, which infect an estimated 511 and 412 million people, respectively5, as well as the hookworms Necator americanus, Ancylostoma duodenale and Ancylostoma ceylanicum, which collectively infect up to 186 million people globally5. Infected individuals can suffer from severe abdominal discomfort, bloody diarrhoea, stunted development and anaemia. Other examples include the filarial nematodes Wuchereria bancrofti and Brugia malayi, the causative agents of lymphatic filariasis that infect up to 96 million people globally5,6, and Onchocerca volvulus, which infects up to 21 million people5. In the case of O. volvulus, recent estimates indicate that 14.6 million are afflicted with skin disease and 1.15 million with blindness7. Furthermore, there has been increasing recognition of a disease known as onchocerciasis-associated epilepsy (OAE), occurring in children and adolescents in onchocerciasis meso- and hyperendemic foci across sub-Saharan Africa8. This condition manifests as a variety of epileptic seizures, including uncontrollable repeated head nodding (‘nodding syndrome’), as well as severe stunting, delayed puberty and impaired mental health (Nakalanga syndrome)9. OAE has been epidemiologically linked to infection with O. volvulus10, but the pathogenesis has yet to be identified8.
A variety of viruses can be found infecting several human parasitic protozoa, including Plasmodium vivax, Trichomonas vaginalis and Cryptosporidium parvum11,12. Viruses infecting Leishmania sp. have been studied in great detail13 and can increase disease severity, parasite prevalence and potentially the incidence rates of both drug resistance and mucocutaneous leishmaniasis14,15. RNA virus infections have been identified in plant-parasitic nematodes16, parasitic flatworms17,18 and free-living nematodes17,19,20, although the impact of viral infections on the biology of the worms is largely unknown.
Here we analysed the transcriptomes of 41 parasitic nematode species infecting humans and animals and discovered 91 virus or virus-like genomic sequences across 28 species. We further characterize the viruses infecting B. malayi and O. volvulus, describing their genomic diversity, geographic spread, phylogeny, abundance throughout different developmental stages, tissue tropism, localization and vertebrate host serology. Finally, we show that an RNA interference (RNAi) response is induced in B. malayi against BMRV1, providing evidence for active viral replication.
[…]
Discussion
We reveal an abundant and diverse RNA virome spanning 14 different viral orders and 24 families within parasitic nematodes. Of the 91 viruses discovered, only 5 have been previously reported, including 3 from A. suum and A. lumbricoides23,25. Our survey is probably an under-representation of the true extent and diversity of the parasitic nematode RNA virome owing to a variety of factors including variations in sample preparation resulting in discarded viral reads and the restricted number, or lack, of transcriptomes for several important parasites. Nevertheless, our analysis supports a conserved global spread of virus–nematode associations across multiple continents in the case of the viruses of A. suum and A. lumbricoides, and O. volvulus, suggesting an ancient and stable co-evolution. This is perhaps best exemplified by members of the Trichinellidae (Supplementary Fig. 1), which show a close evolutionary relationship, as well as phylogenetic clustering of diverse virus sequences from different species and orders of parasitic nematodes.
The parasitic nematodes identified with viruses include several important human parasitic nematodes, A. lumbricoides, T. trichiura, O. volvulus, B. malayi, A. ceylanicum and Trichinella spiralis, which cause substantial public health issues, with over 1.5 billion people infected with one or more such parasites4,5,6,44,45. Several other species cause an even greater global burden in the livestock industry46, with 15 economically important parasites (A. suum, Dictyocaulus viviparous, Haemonchus contortus, Ostertagia ostertagi, Oesophagostomum dentatum, Teladorsagia circumcinta, Trichuris suis plus 8 Trichinella spp.) of cattle, sheep and pigs, harbouring 37 previously unreported viruses.
The full extent and diversity of the parasitic nematode RNA virome, how it impacts nematode biology and whether they act as drivers or modulators of disease pathogenesis remain critical knowledge gaps. Indeed, in the parasite Toxocara canis, which causes neurotoxocariasis, components of the TCLA virus have been reported to be highly expressed in infective larvae (18% of expressed sequence tags) before entry into a vertebrate host (for example, humans and dogs)29, with human infections eliciting antibody responses against several TCLA virus proteins29, indicating potential roles in transmission and infectivity. Alternatively, extrapolation from the most well-characterized RNA viruses of Leishmania sp. protozoan parasites suggests potential roles of nematode viruses in disease pathology and progression. Both Leishmania virus 1 (LRV1) and T. vaginalis virus induce hyperinflammatory immunity, which drives disease pathogenesis and subverts host immunity to the parasites’ advantage14,15,47. We show that BMRV1 and OVRV1 elicit antibody responses from the host showing direct exposure to the immune system, and we speculate that this suggests the potential to modulate host immunity to the parasite and cross-reactive immunity to other RNA viruses.
[…]
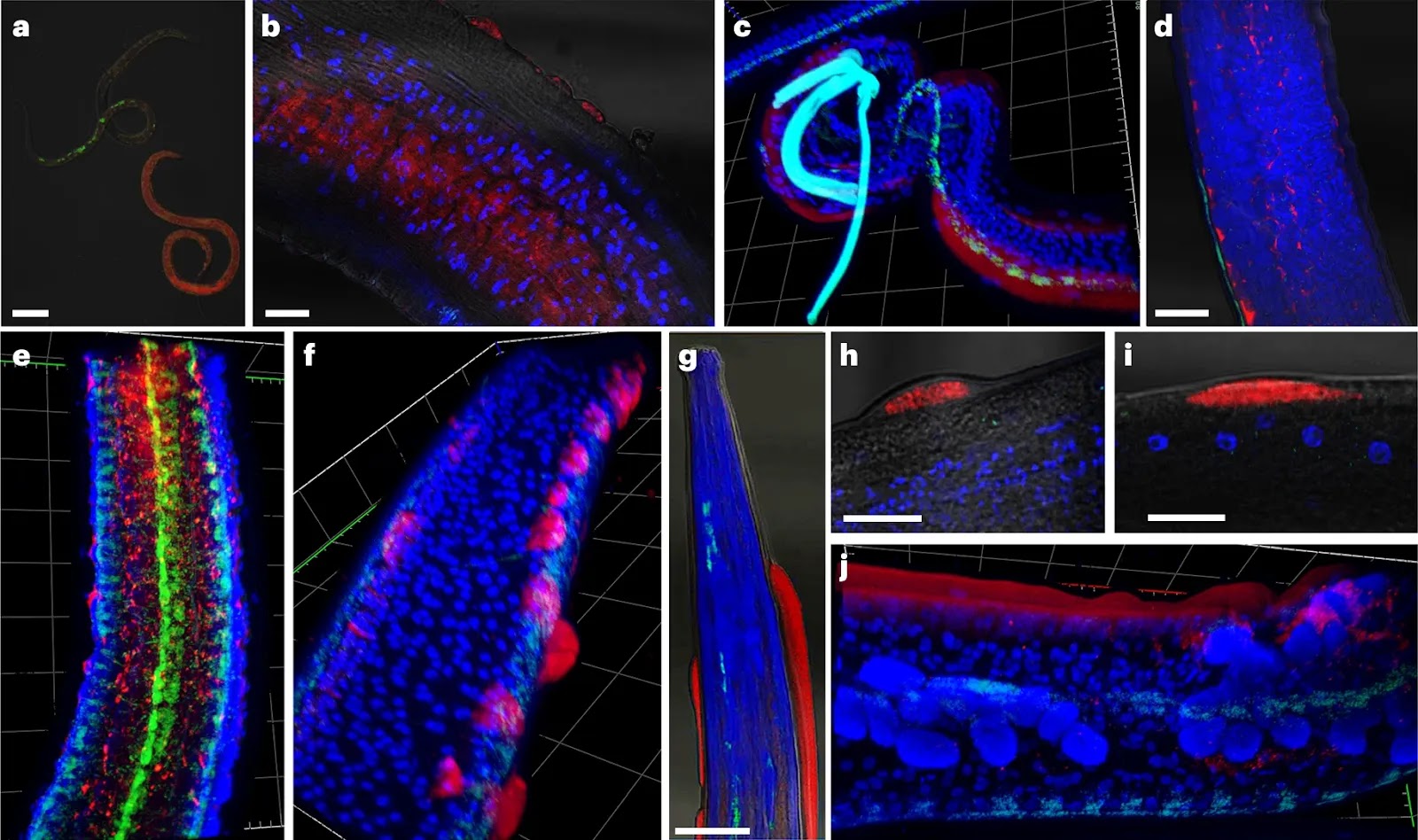
Fig. 4: Representative FISH microscopy images of B. malayi showing localization of virus RNA within nematode tissues, alongside the Wolbachia endosymbiont as a technical control.
Virus RNA stained red; Wolbachia stained green; DAPI nuclear stain blue. a–e, Note the different levels of viral infection in microfilariae (a), localization of the viral stain in male testes (b) and the hypodermal cells near the male spicule (c). Virus signal within adult female reproductive tracts appears between developing eggs within the paired uteri of adult females, with early embrys in the left uteri and ‘pretzel-stage’ microfilariae in the right (d), with the developing eggs casting a ‘shadow’ in between virus staining, visible in 3D images of female uteri (e). f–j, In older adults (>12 months), we observed ‘epicuticular inflations’ often with an intense viral signal (f), typically occurring near the head (g) or tail regions of the nematodes. They can appear as single separate inflations at different nematode orientations, either next to internal organs (h) or the hypodermal chords (i), or as a continuous inflation along the nematode flank (j). Scale bars measure 20 µm (a,b,h,i) or 50 µm (d,g). Gridlines in three-dimensional 𝓏-stack figures (c,e,f,j) measure 40 µm by 40 µm. A total of 15 adult male and female parasites were processed in separate experiments. Parasites with epicuticular inflations were typically between 12 and 19 months at the time of sampling, with jird animal hosts being 15–22 months of age, respectively. Parasites without were typically 3–6 months of age, with the jird animal hosts being 6–9 months of age.
Fig. 5: Validation of OVRV1 using RT-PCR, western blot and representative IFA staining of O. volvulus nodules with anti-OVRV1 glycoprotein antibodies.
Anti-OVRV1 glycoprotein antibodies stained green; DAPI nuclear stain blue. a, RT-PCR experiments show that OVRV1 can be amplified only from reverse-transcribed RNA, from both O. volvulus (lane 1, n = 1) and O. ochengi (lanes 2–4, n = 3). b, Western blots against the OVRV1 glycoprotein show different molecular weight bands occurring depending on the life cycle stage of O. volvulus (n = 3). All IFA images include the DAPI nuclear stain (blue). c,d, Images of the paired uteri from adult O. volvulus females show virus stains surrounding and entering developing embryos within the uteri (solid arrow), while surrounding but not within the early embryos (hollow arrow). Developing embryos can show either complete infection rates (c) or a much smaller proportion (d). e, Mature microfilariae released from the female, located within surrounding nodule tissues, stain heavily for OVRV1 glycoprotein. f,g, Intense antibody staining is observed surrounding the nematode rachis, where eggs are first formed (solid arrows). The heavily stained rachis is either surrounded by early-stage eggs with green staining surrounding them (f) or without surrounding eggs (g). h,i, Cellular inflations containing intense antibody staining are observed on the external face of the adult female uterine walls (solid arrows). j,k, Male O. volvulus are frequently observed to be infected, with viral stains occurring in different tissues (j), as well as surrounding and entering the male testes (k). Parasites were obtained from sections of fixed O. volvulus nodules from human patients (n = 8 nodules).
So, if you reject the evolutionary explanation of these viruses-nematode associations in favour of a creationist 'intelligent [sic] design' explanation you must assume the designer intended the consequences of its design since it is axiomatic of the creationist cult that the designer is a perfect, omniscient god for whom the consequences of its design must have been known in advance and so were designed with that function in mind.
And of course we can dismiss the childish nonsense about 'genetic entropy; causing 'devolution' [sic] because these viruses are clearly gaining an advantage in infecting the nematodes because that gives them easy access to their vertebrate hosts, and anything which conveys an advantage is evolution, not 'devolution'. Only someone ignorant of evolution would fall for such biologically nonsensical excuse for parasites, as any biologist worthy of the term would have known before he came up with it.
So, the question remains unanswered by creationists - is this an example of malevolent design, or of evolution?
Heliconius erato with one wing (right) altered by CRISPR gene. Dorsal view
Luca Livraghi.
Heliconius erato with one wing (right) altered by CRISPR gene. Ventral view
Luca Livraghi.
This is one of those research papers that creationists traditionally misrepresent as 'proving' that something biologists have long known about is wrong, and so it somehow discredits the entire body of science, and, in this specific case, that the existence of RNA that is non-coding but functional, somehow proves there is no such thing as 'junk' DNA.
In reality, of course, it simply shows us that sometimes, evolution happens in unexpected ways.
The assumption, which is almost always valid and based on sound scientific evidence, is that DNA is transcribed into messenger RNA (mRNA) that mRNA is transcribed by ribosomes into proteins. Proteins then serve functions such as structural proteins or as enzymes catalysing chemical processes in the cell. In the special case of so-called homeobox genes (hox genes) these have a regulatory function in a developing embryo, stimulating groups of cells to organise into specific tissues such as limbs, eyes and other specialised tissues, by switching other genes off and on at certain times.
It can only be by remaining wilfully ignorant of the history of life on Earth and how it has been affected by catastrohic changes, that creationists are able to maintain the childish delusion that Earth is somehow fine-tuned for life, and especially their life.
The fact that very much of the surface of Earth, such as oceans, deserts and high mountain ranges is lethal without special equipment, and without cloths, shelter and fire , most of the northern and southern latitudes are not survivable in an average winter should be a clue to the fallacy of the claim, but then, from the safety of small town, Bible Belt America, where the rest of the world might as well not exist, such places can be safely ignored.
But the fact remains, whether ignored or not, that there have been several periods in the history of life on Earth that the planet has become a very hostile place, resulting in mass extinctions wheree life looked as though someone had hit the reset button. The primary cause of these fluctuations is almost certinly plate techtonics which, through mid-ocean ridge activity, release sulphates and phosphates which produce blooms of plankton which then die, locking up carbon and producing anoxic sediments on the ocean bed, and by releasing CO2 into the atmosphere where if causes a rise in ocean temperature.
Volcanic activity also produces basalt which, when eroded and washed into the sea adds to the phosphates which produce planktonic blooms and subsequent deoxygenation.
How these terrestrial and marine sources act together to produce the frequent OAEs has now been explained.